

Probing excited state 1Hα chemical shifts in intrinsically disordered proteins with a triple resonance-based CEST experiment: Application to a disorder-to-order switch
- PMID: 37607621
- PMCID: PMC7615522
- DOI: 10.1016/j.ymeth.2023.08.009
Probing excited state 1Hα chemical shifts in intrinsically disordered proteins with a triple resonance-based CEST experiment: Application to a disorder-to-order switch
Abstract
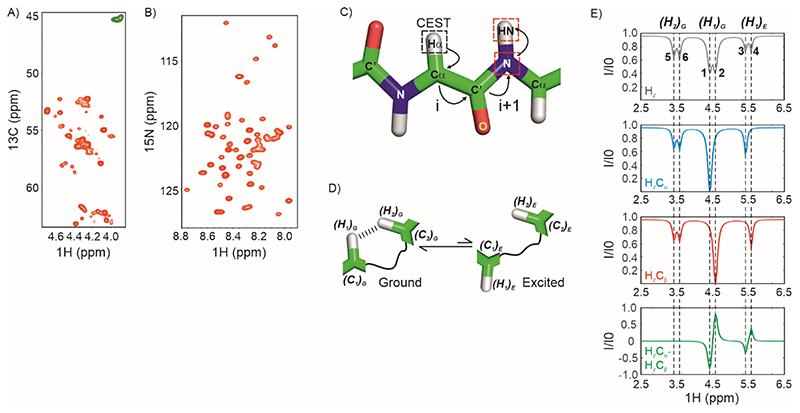
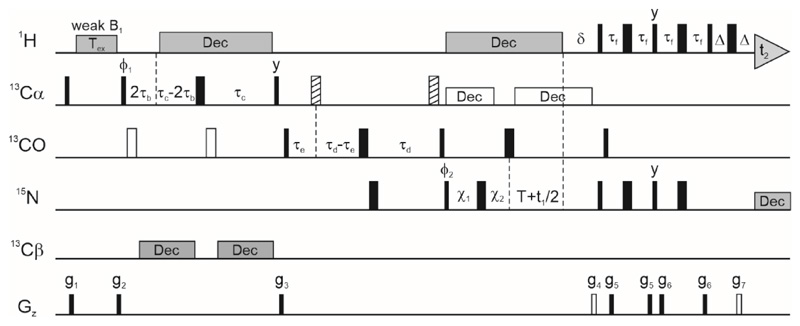
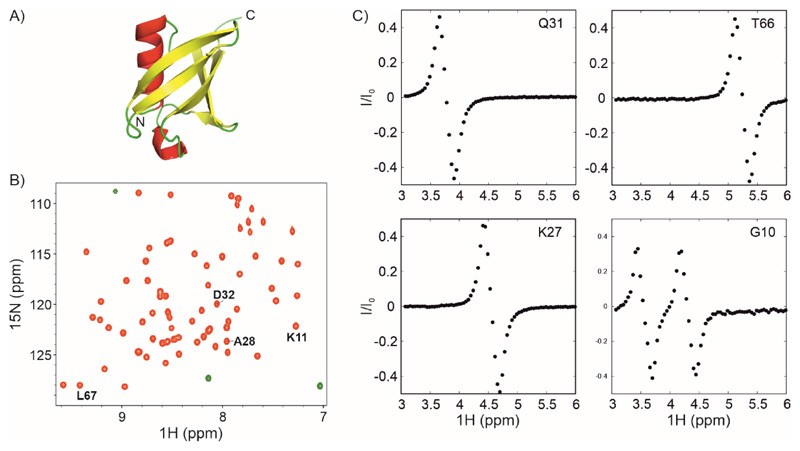
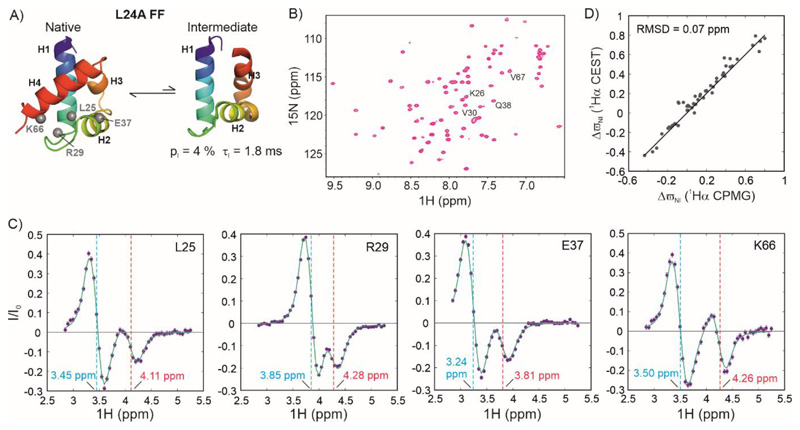
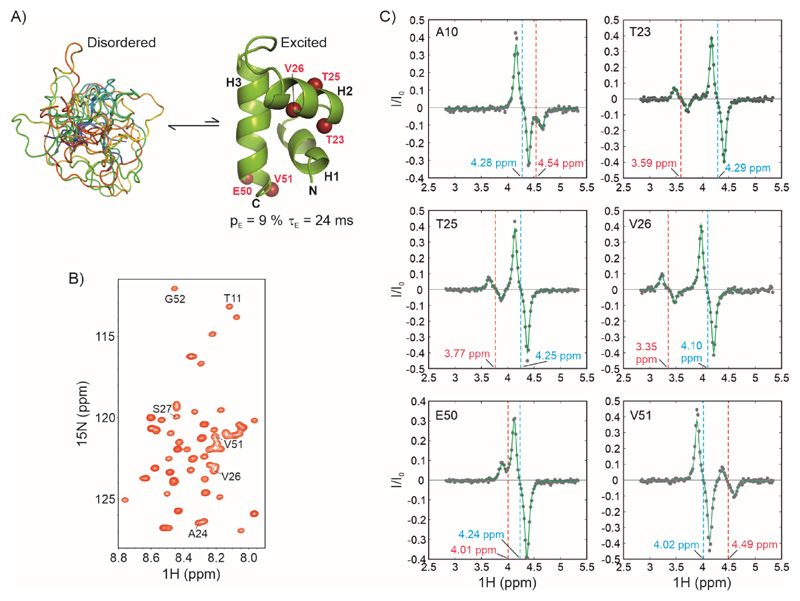
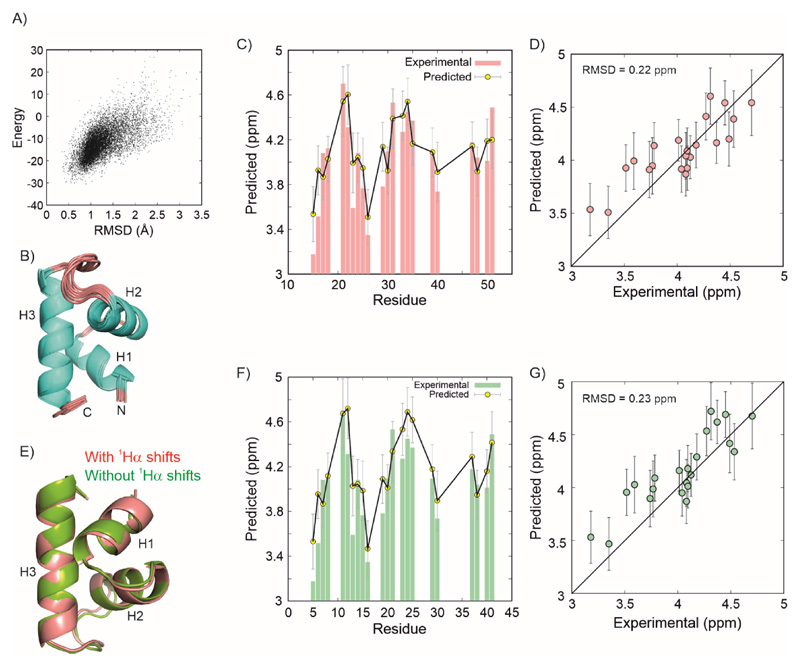
Over 40% of eukaryotic proteomes and 15% of bacterial proteomes are predicted to be intrinsically disordered based on their amino acid sequence. Intrinsically disordered proteins (IDPs) exist as heterogeneous ensembles of interconverting conformations and pose a challenge to the structure-function paradigm by apparently functioning without possessing stable structural elements. IDPs play a prominent role in biological processes involving extensive intermolecular interaction networks and their inherently dynamic nature facilitates their promiscuous interaction with multiple structurally diverse partner molecules. NMR spectroscopy has made pivotal contributions to our understanding of IDPs because of its unique ability to characterize heterogeneity at atomic resolution. NMR methods such as Chemical Exchange Saturation Transfer (CEST) and relaxation dispersion have enabled the detection of 'invisible' excited states in biomolecules which are transiently and sparsely populated, yet central for function. Here, we develop a 1Hα CEST pulse sequence which overcomes the resonance overlap problem in the 1Hα-13Cα plane of IDPs by taking advantage of the superior resolution in the 1H-15N correlation spectrum. In this sequence, magnetization is transferred after 1H CEST using a triple resonance coherence transfer pathway from 1Hα (i) to 1HN(i + 1) during which the 15N(t1) and 1HN(t2) are frequency labelled. This approach is integrated with spin state-selective CEST for eliminating spurious dips in CEST profiles resulting from dipolar cross-relaxation. We apply this sequence to determine the excited state 1Hα chemical shifts of the intrinsically disordered DNA binding domain (CytRN) of the bacterial cytidine repressor (CytR), which transiently acquires a functional globally folded conformation. The structure of the excited state, calculated using 1Hα chemical shifts in conjunction with other excited state NMR restraints, is a three-helix bundle incorporating a helix-turn-helix motif that is vital for binding DNA.
Keywords: Alpha proton; Chemical exchange saturation transfer; Conformational change; Intrinsically disordered proteins; Nuclear magnetic resonance spectroscopy; Protein dynamics.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
- Uversky VN. Intrinsically disordered proteins and their “mysterious”(meta) physics. Front Phys. 2019;7:10.
-
- Gianni S, Dogan J, Jemth P. Coupled binding and folding of intrinsically disordered proteins: what can we learn from kinetics? Curr Opin Struct Biol. 2016;36:18. - PubMed
-
- Uversky VN. Unusual biophysics of intrinsically disordered proteins. Biochim Biophys Acta: Proteins Proteomics. 2013;1834:932. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
