

This is a preprint.
Machine-guided design of synthetic cell type-specific cis-regulatory elements
- PMID: 37609287
- PMCID: PMC10441439
- DOI: 10.1101/2023.08.08.552077
Machine-guided design of synthetic cell type-specific cis-regulatory elements
Update in
-
Machine-guided design of cell-type-targeting cis-regulatory elements.Nature. 2024 Oct;634(8036):1211-1220. doi: 10.1038/s41586-024-08070-z. Epub 2024 Oct 23. Nature. 2024. PMID: 39443793 Free PMC article.
Abstract
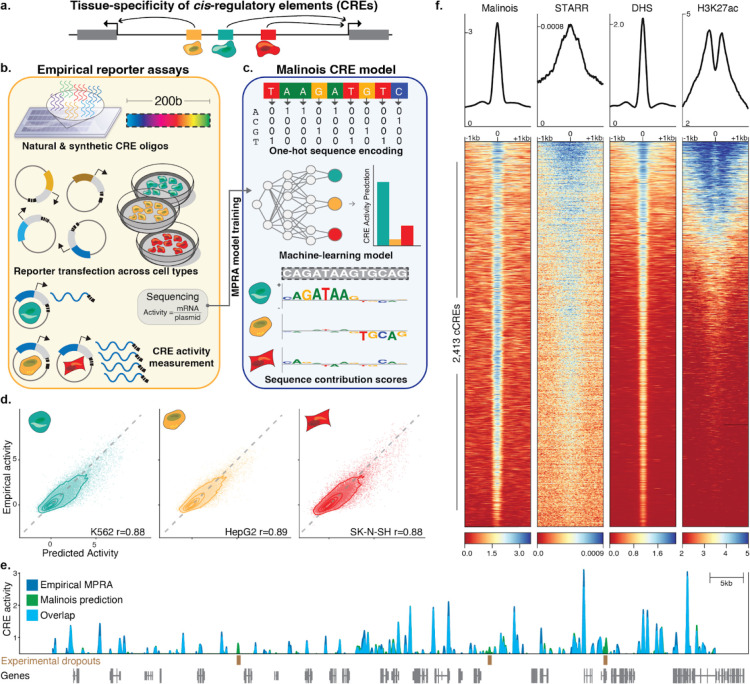
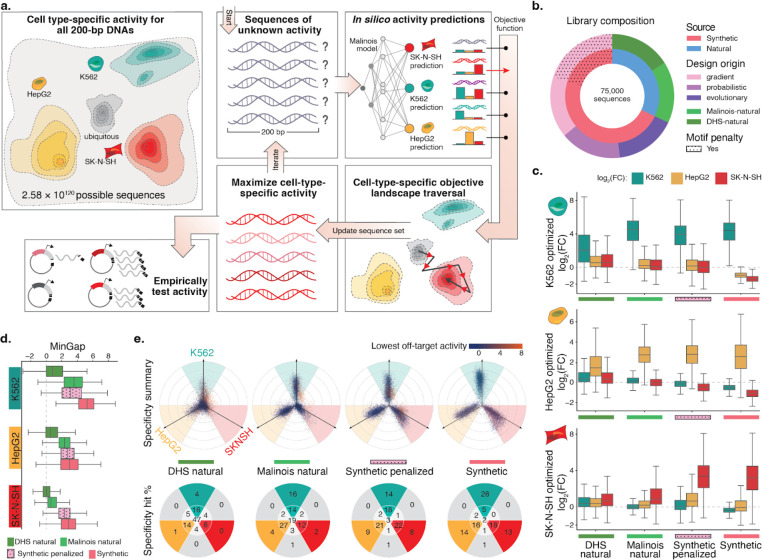
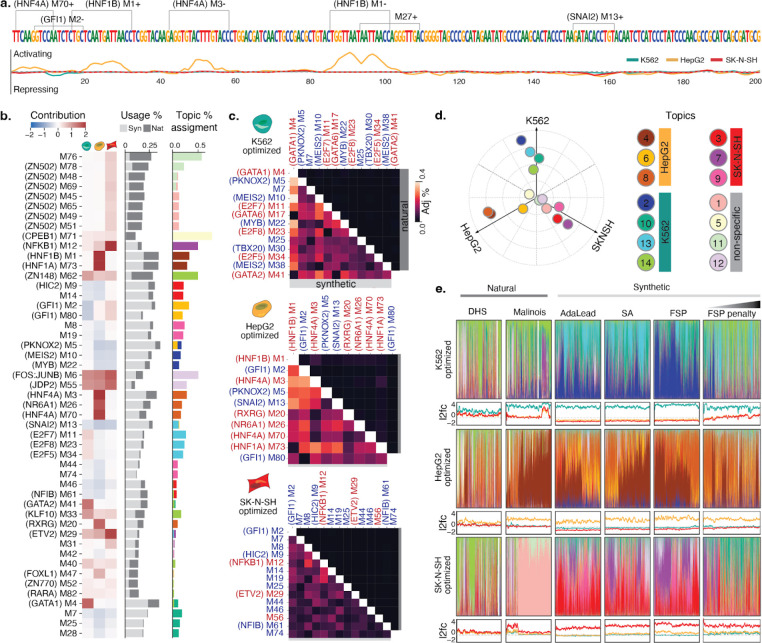
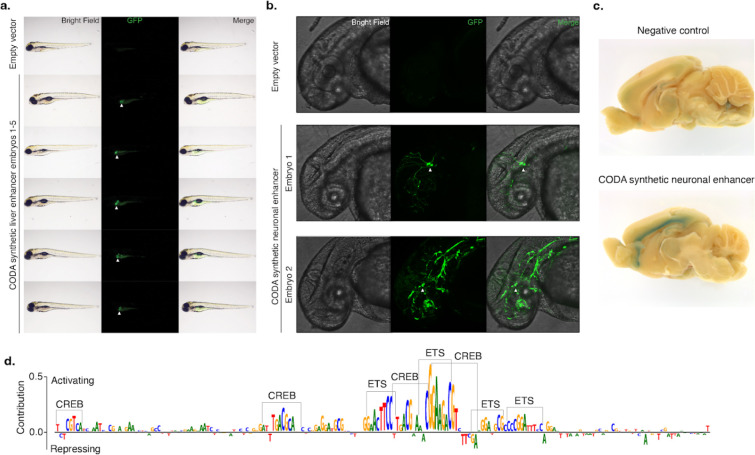
Cis-regulatory elements (CREs) control gene expression, orchestrating tissue identity, developmental timing, and stimulus responses, which collectively define the thousands of unique cell types in the body. While there is great potential for strategically incorporating CREs in therapeutic or biotechnology applications that require tissue specificity, there is no guarantee that an optimal CRE for an intended purpose has arisen naturally through evolution. Here, we present a platform to engineer and validate synthetic CREs capable of driving gene expression with programmed cell type specificity. We leverage innovations in deep neural network modeling of CRE activity across three cell types, efficient in silico optimization, and massively parallel reporter assays (MPRAs) to design and empirically test thousands of CREs. Through in vitro and in vivo validation, we show that synthetic sequences outperform natural sequences from the human genome in driving cell type-specific expression. Synthetic sequences leverage unique sequence syntax to promote activity in the on-target cell type and simultaneously reduce activity in off-target cells. Together, we provide a generalizable framework to prospectively engineer CREs and demonstrate the required literacy to write regulatory code that is fit-for-purpose in vivo across vertebrates.
Conflict of interest statement
Competing Interests PCS is a co-founder of and consultant to Sherlock Biosciences and Board Member of Danaher Corporation. PCS and RT have filed intellectual property related to MPRA. SJG, RIC, SKR, PCS, and RT have filed a provisional patent application related to work described here.
Figures
References
-
- Wittkopp P. J. & Kalay G. Cis-regulatory elements: molecular mechanisms and evolutionary processes underlying divergence. Nat. Rev. Genet. 13, 59–69 (2011). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials