

Structural insights of the p97/VCP AAA+ ATPase: How adapter interactions coordinate diverse cellular functionality
- PMID: 37611827
- PMCID: PMC10641518
- DOI: 10.1016/j.jbc.2023.105182
Structural insights of the p97/VCP AAA+ ATPase: How adapter interactions coordinate diverse cellular functionality
Abstract
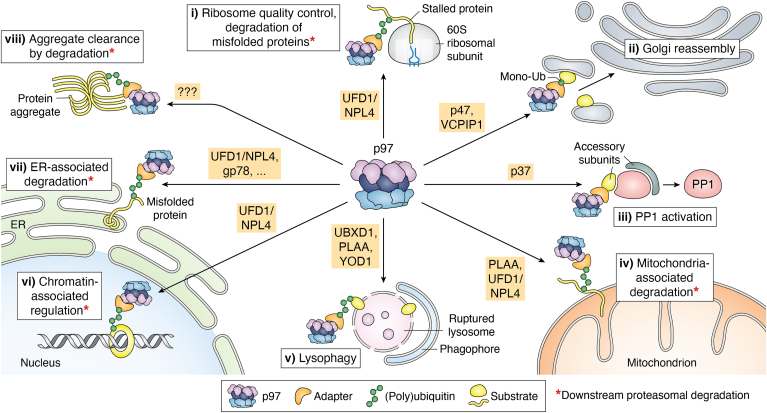
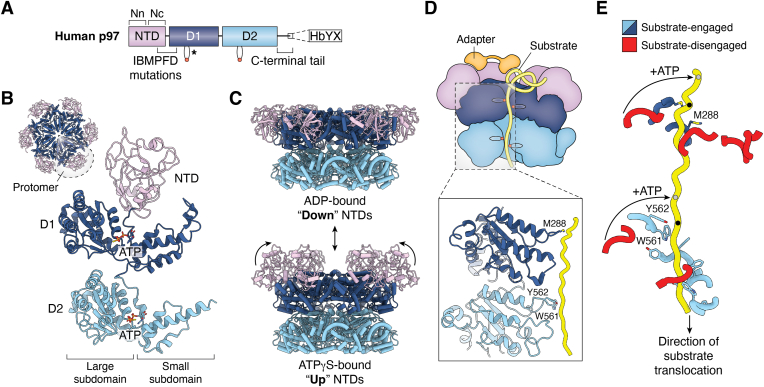
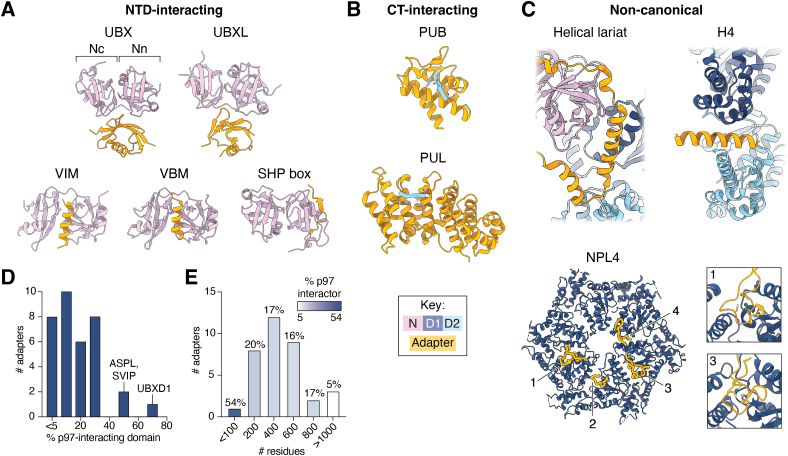
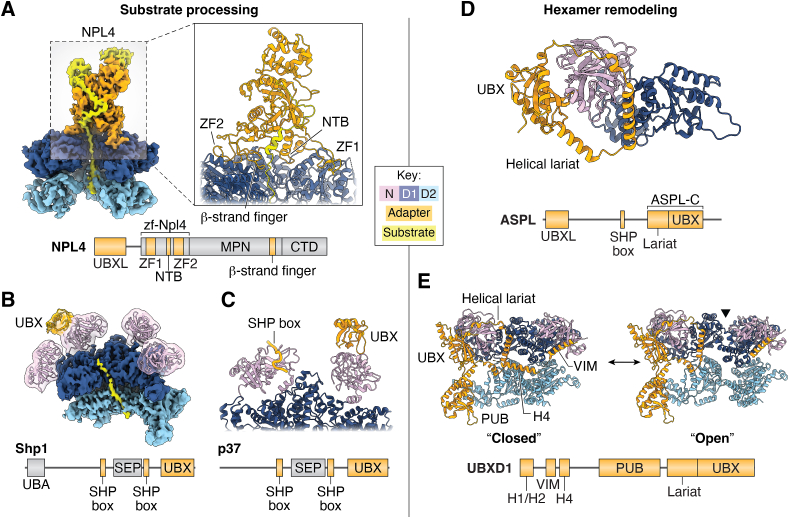
p97/valosin-containing protein is an essential eukaryotic AAA+ ATPase with diverse functions including protein homeostasis, membrane remodeling, and chromatin regulation. Dysregulation of p97 function causes severe neurodegenerative disease and is associated with cancer, making this protein a significant therapeutic target. p97 extracts polypeptide substrates from macromolecular assemblies by hydrolysis-driven translocation through its central pore. Growing evidence indicates that this activity is highly coordinated by "adapter" partner proteins, of which more than 30 have been identified and are commonly described to facilitate translocation through substrate recruitment or modification. In so doing, these adapters enable critical p97-dependent functions such as extraction of misfolded proteins from the endoplasmic reticulum or mitochondria, and are likely the reason for the extreme functional diversity of p97 relative to other AAA+ translocases. Here, we review the known functions of adapter proteins and highlight recent structural and biochemical advances that have begun to reveal the diverse molecular bases for adapter-mediated regulation of p97 function. These studies suggest that the range of mechanisms by which p97 activity is controlled is vastly underexplored with significant advances possible for understanding p97 regulation by the most known adapters.
Keywords: AAA+ ATPase; AlphaFold; ERAD; VCP; adapter; adaptor; autophagy; cofactor; cryo-EM; molecular chaperone; p97; protein structure prediction; proteostasis; unfoldase.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
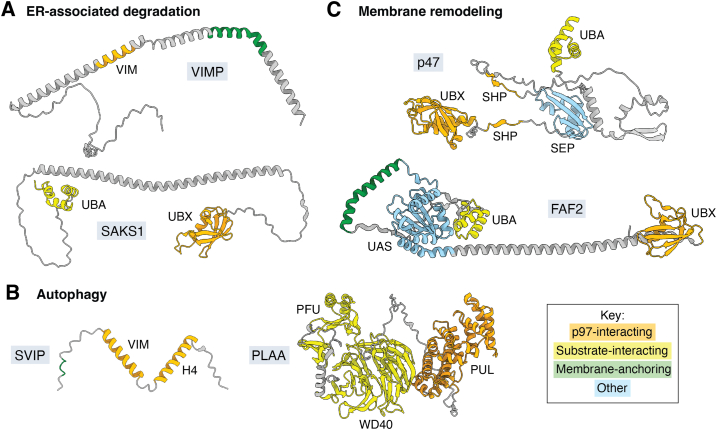
References
-
- Prakash S., Tian L., Ratliff K.S., Lehotzky R.E., Matouschek A. An unstructured initiation site is required for efficient proteasome-mediated degradation. Nat. Struct. Mol. Biol. 2004;11:830–837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
