

A novel recombination protein C12ORF40/REDIC1 is required for meiotic crossover formation
- PMID: 37612290
- PMCID: PMC10447524
- DOI: 10.1038/s41421-023-00577-5
A novel recombination protein C12ORF40/REDIC1 is required for meiotic crossover formation
Abstract
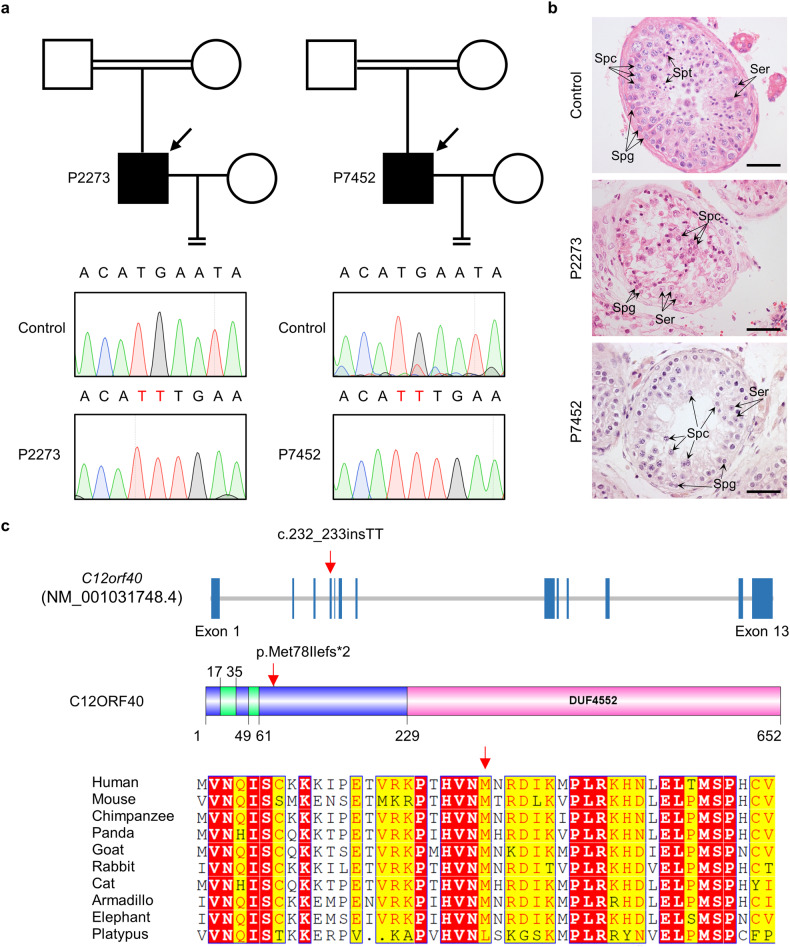
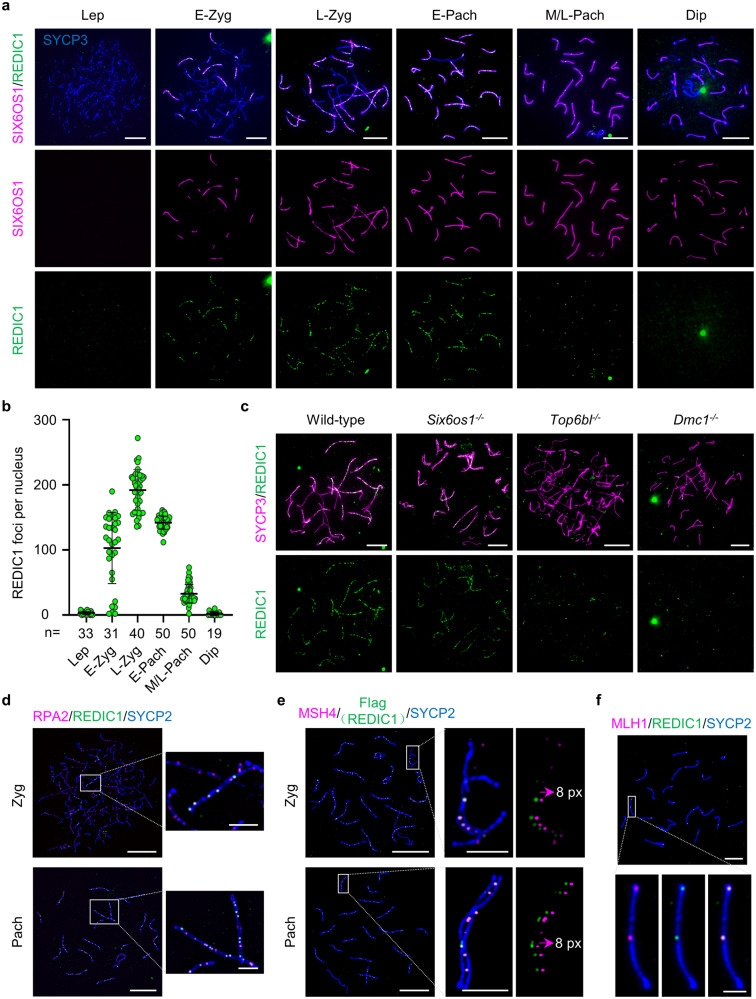
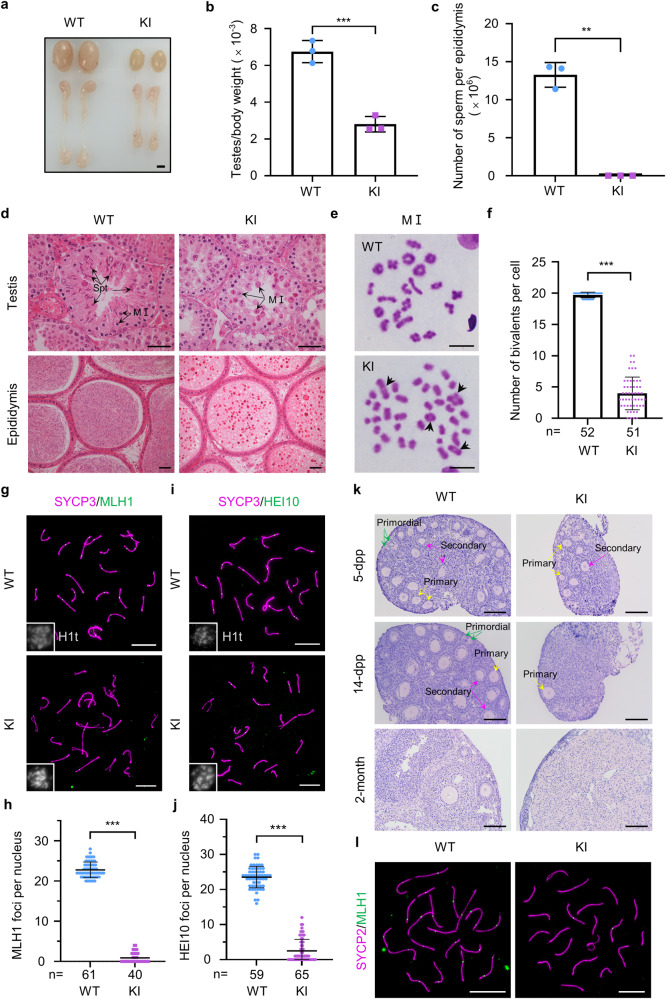
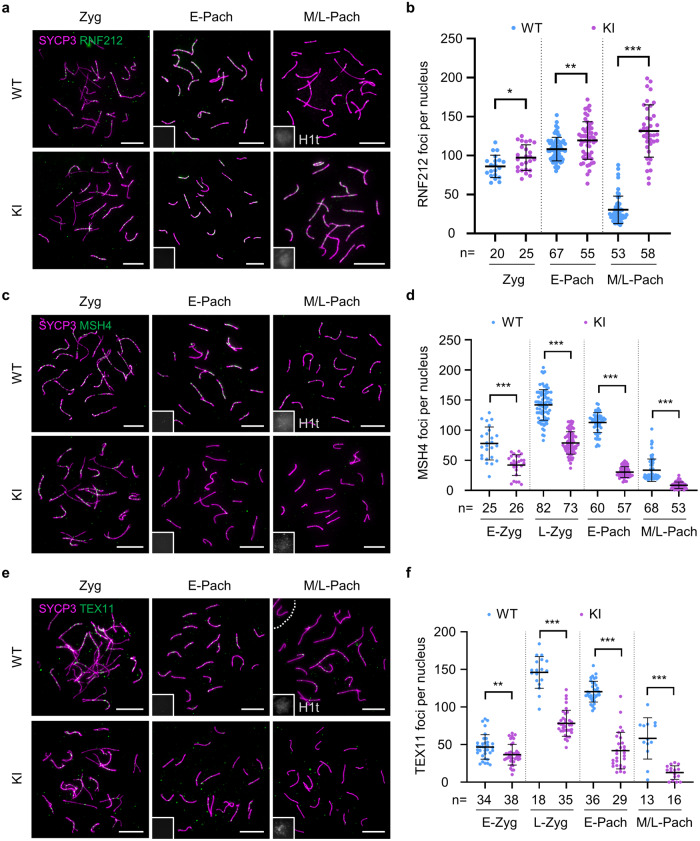
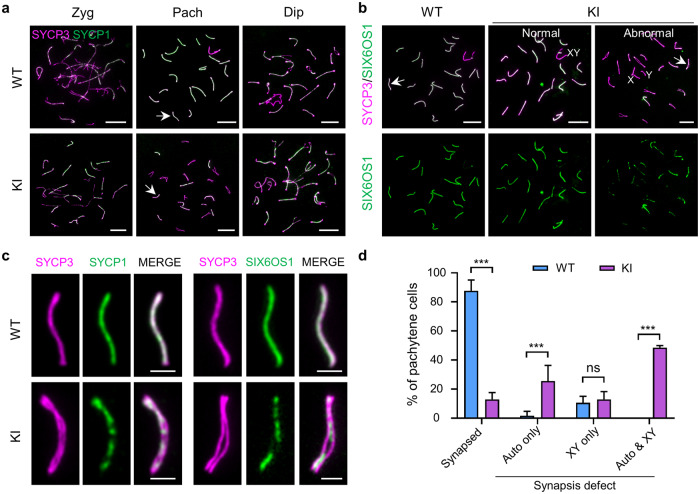
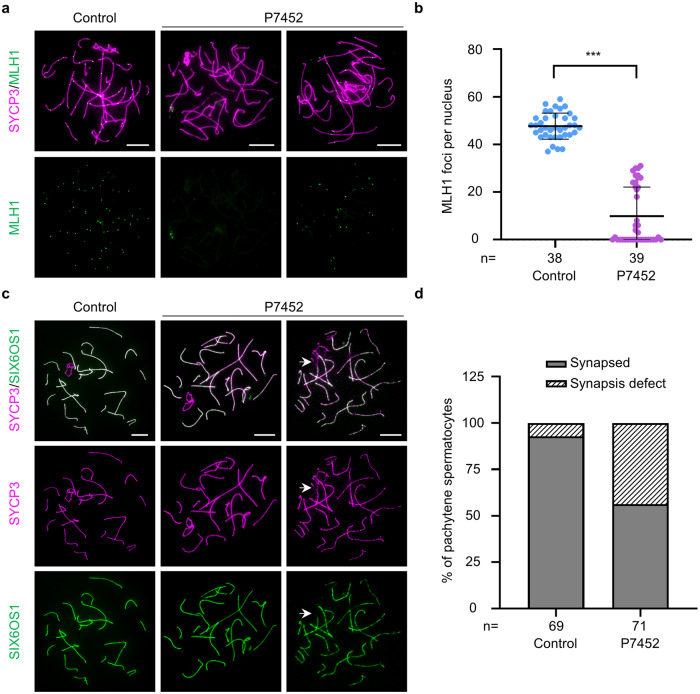
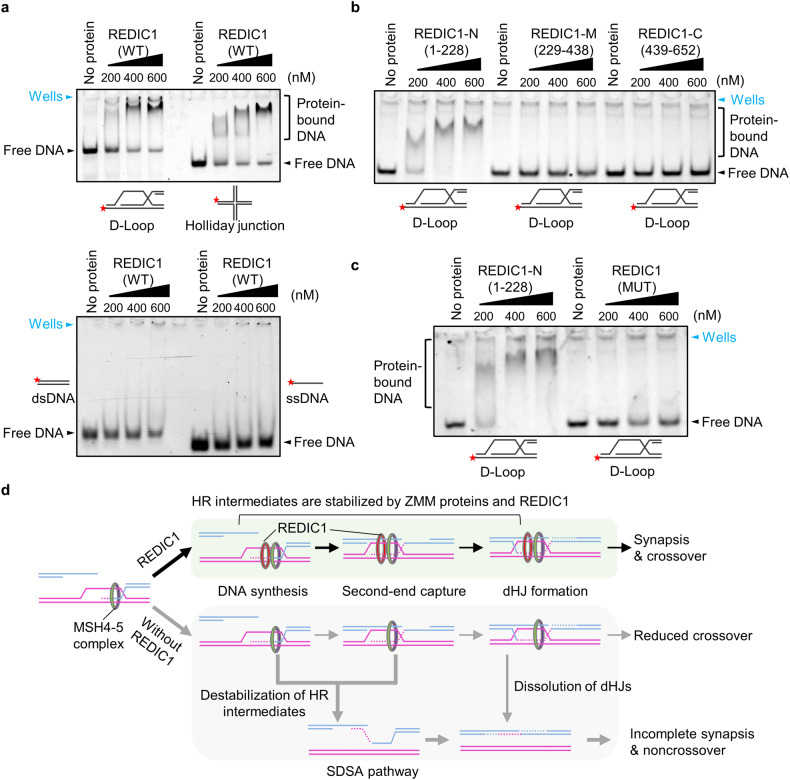
During meiosis, at least one crossover must occur per homologous chromosome pair to ensure normal progression of meiotic division and accurate chromosome segregation. However, the mechanism of crossover formation is not fully understood. Here, we report a novel recombination protein, C12ORF40/REDIC1, essential for meiotic crossover formation in mammals. A homozygous frameshift mutation in C12orf40 (c.232_233insTT, p.Met78Ilefs*2) was identified in two infertile men with meiotic arrest. Spread mouse spermatocyte fluorescence immunostaining showed that REDIC1 forms discrete foci between the paired regions of homologous chromosomes depending on strand invasion and colocalizes with MSH4 and later with MLH1 at the crossover sites. Redic1 knock-in (KI) mice homozygous for mutation c.232_233insTT are infertile in both sexes due to insufficient crossovers and consequent meiotic arrest, which is also observed in our patients. The foci of MSH4 and TEX11, markers of recombination intermediates, are significantly reduced numerically in the spermatocytes of Redic1 KI mice. More importantly, our biochemical results show that the N-terminus of REDIC1 binds branched DNAs present in recombination intermediates, while the identified mutation impairs this interaction. Thus, our findings reveal a crucial role for C12ORF40/REDIC1 in meiotic crossover formation by stabilizing the recombination intermediates, providing prospective molecular targets for the clinical diagnosis and therapy of infertility.
© 2023. Center for Excellence in Molecular Cell Science, CAS.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Page SL, Hawley RS. Chromosome choreography: the meiotic ballet. Science. 2003;301:785–789. - PubMed
Grants and funding
- 31890780/National Natural Science Foundation of China (National Science Foundation of China)
- 32061143006/National Natural Science Foundation of China (National Science Foundation of China)
- U21A20204/National Natural Science Foundation of China (National Science Foundation of China)
- 32100689/National Natural Science Foundation of China (National Science Foundation of China)
- 31900398/National Natural Science Foundation of China (National Science Foundation of China)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
