

A systematic analysis of the human immune response to Plasmodium vivax
- PMID: 37616070
- PMCID: PMC10575735
- DOI: 10.1172/JCI152463
A systematic analysis of the human immune response to Plasmodium vivax
Abstract
BACKGROUNDThe biology of Plasmodium vivax is markedly different from that of P. falciparum; how this shapes the immune response to infection remains unclear. To address this shortfall, we inoculated human volunteers with a clonal field isolate of P. vivax and tracked their response through infection and convalescence.METHODSParticipants were injected intravenously with blood-stage parasites and infection dynamics were tracked in real time by quantitative PCR. Whole blood samples were used for high dimensional protein analysis, RNA sequencing, and cytometry by time of flight, and temporal changes in the host response to P. vivax were quantified by linear regression. Comparative analyses with P. falciparum were then undertaken using analogous data sets derived from prior controlled human malaria infection studies.RESULTSP. vivax rapidly induced a type I inflammatory response that coincided with hallmark features of clinical malaria. This acute-phase response shared remarkable overlap with that induced by P. falciparum but was significantly elevated (at RNA and protein levels), leading to an increased incidence of pyrexia. In contrast, T cell activation and terminal differentiation were significantly increased in volunteers infected with P. falciparum. Heterogeneous CD4+ T cells were found to dominate this adaptive response and phenotypic analysis revealed unexpected features normally associated with cytotoxicity and autoinflammatory disease.CONCLUSIONP. vivax triggers increased systemic interferon signaling (cf P. falciparum), which likely explains its reduced pyrogenic threshold. In contrast, P. falciparum drives T cell activation far in excess of P. vivax, which may partially explain why falciparum malaria more frequently causes severe disease.TRIAL REGISTRATIONClinicalTrials.gov NCT03797989.FUNDINGThe European Union's Horizon 2020 Research and Innovation programme, the Wellcome Trust, and the Royal Society.
Keywords: Immunology; Infectious disease; Malaria.
Conflict of interest statement
Figures
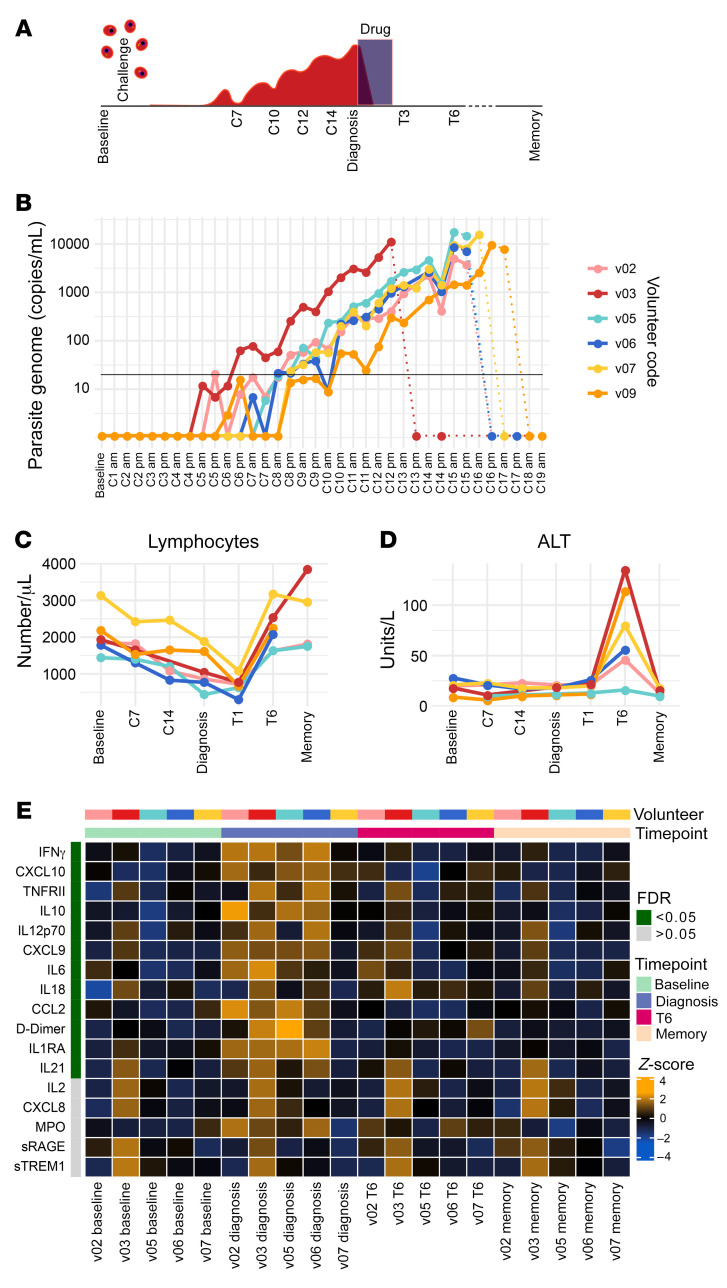
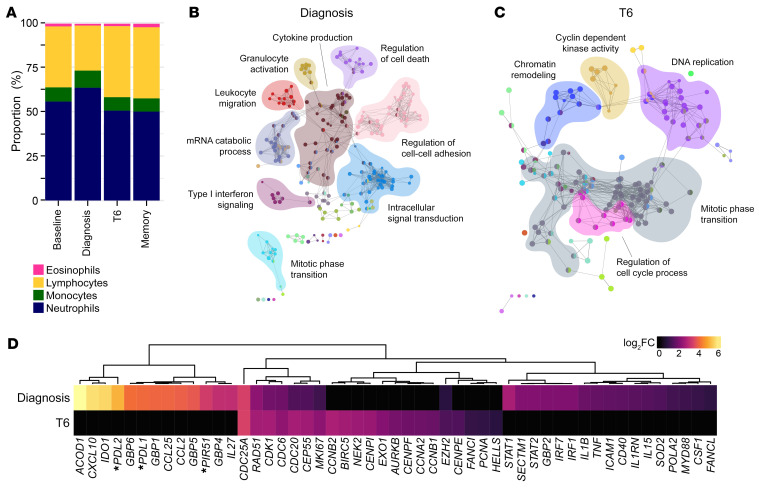
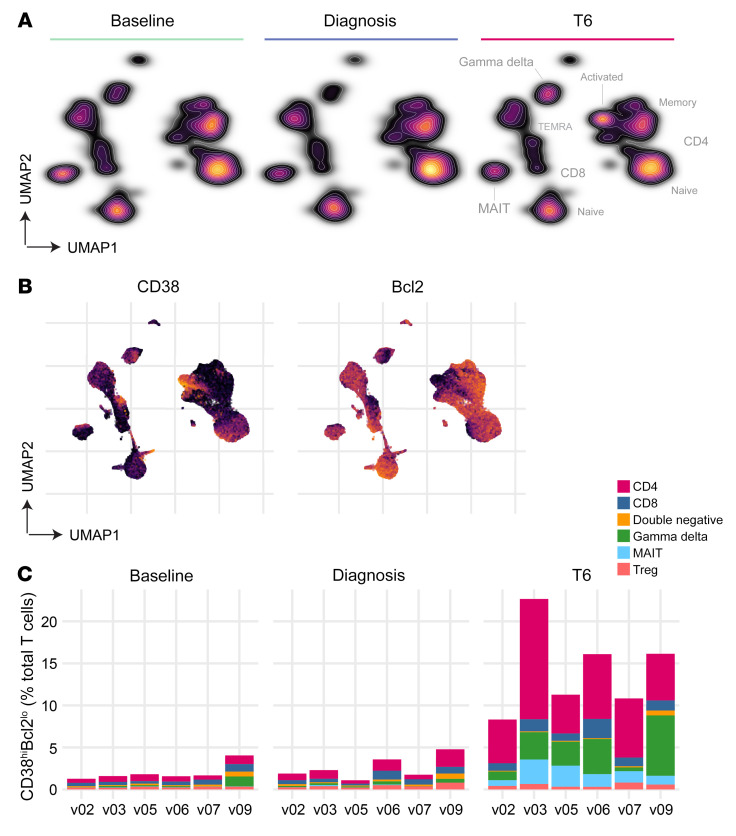
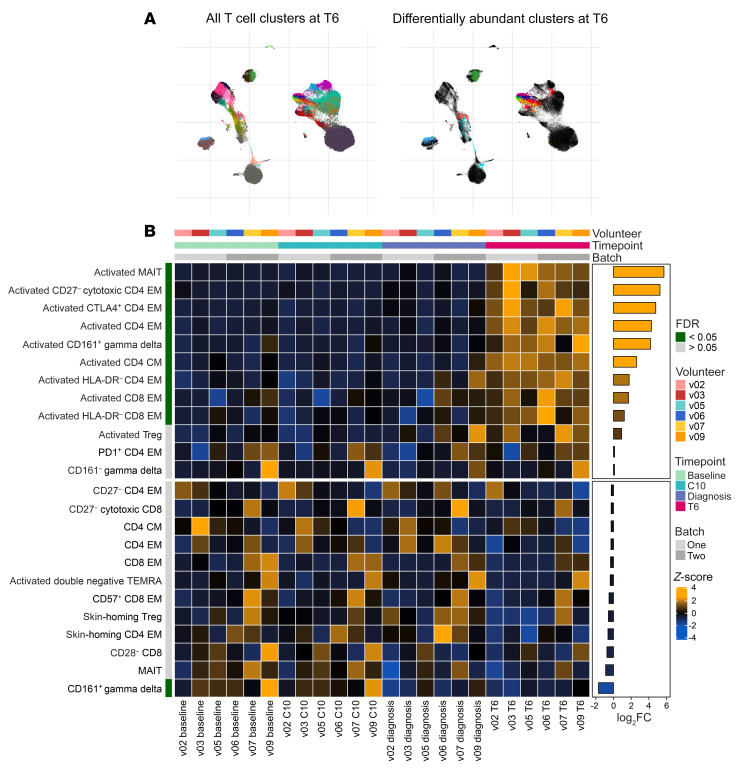
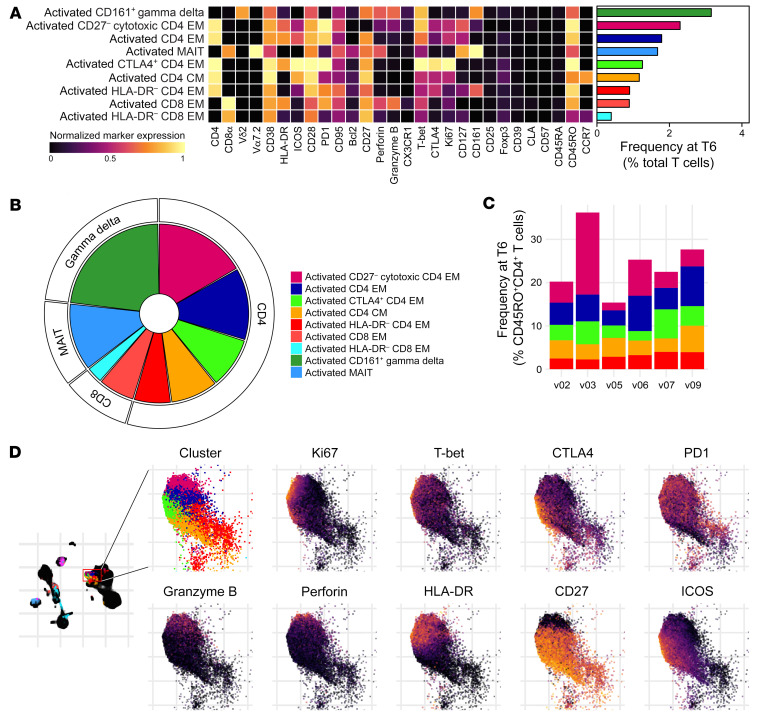
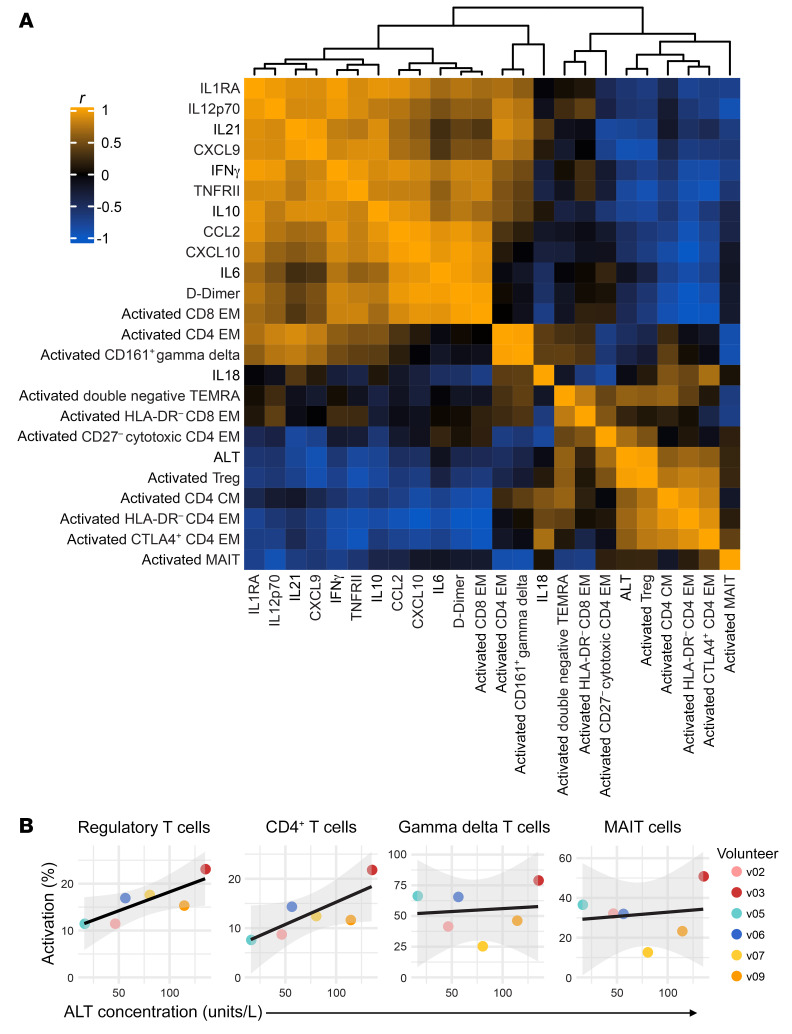
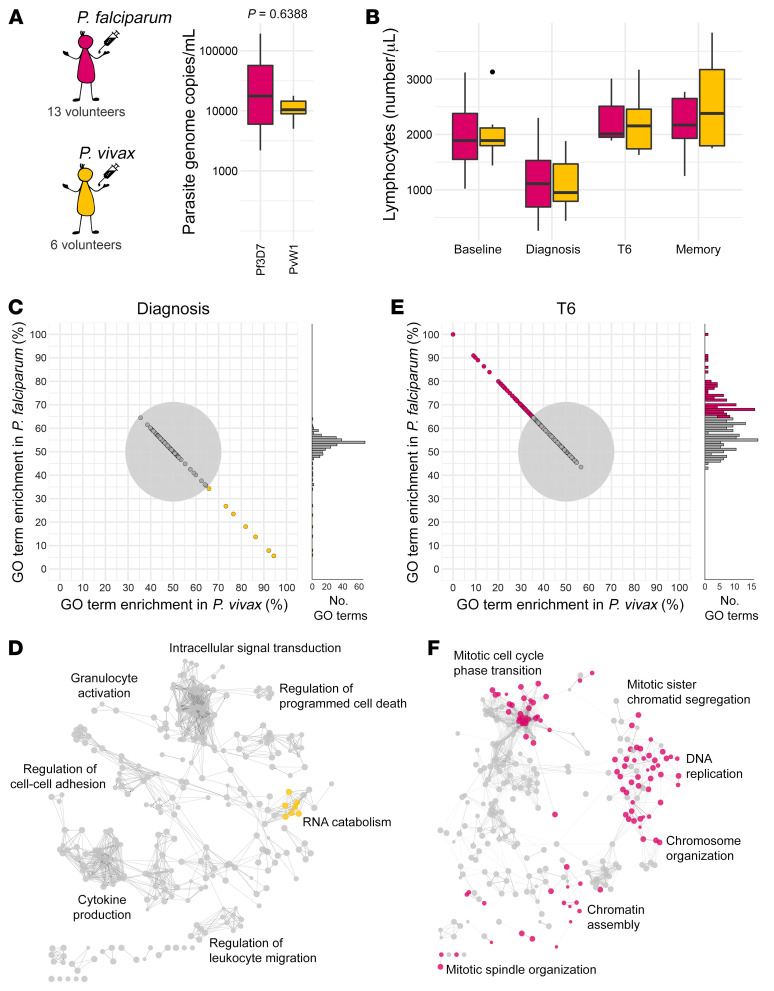
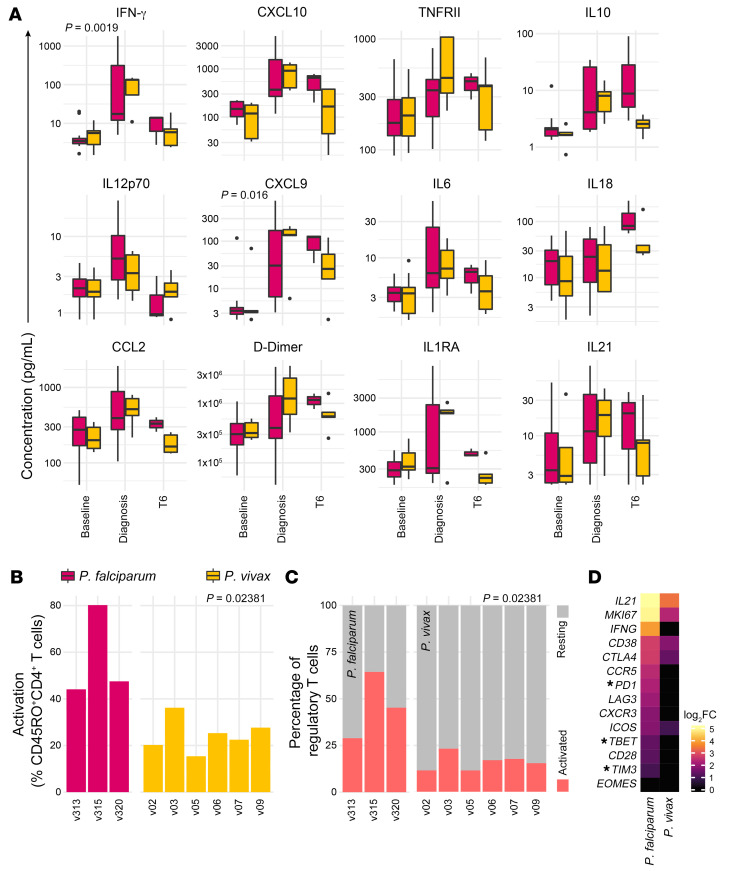
References
Publication types
MeSH terms
Associated data
Grants and funding
- 107668/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- 203764/Z/16/Z/WT_/Wellcome Trust/United Kingdom
- DH_/Department of Health/United Kingdom
- 222323/Z/21/Z/WT_/Wellcome Trust/United Kingdom
- MC_UU_00008/MRC_/Medical Research Council/United Kingdom
- MC_UU_00008/6/MRC_/Medical Research Council/United Kingdom
- 106917/Z/15/Z/WT_/Wellcome Trust/United Kingdom
- WT_/Wellcome Trust/United Kingdom
- 206194/Z/17/Z/WT_/Wellcome Trust/United Kingdom
- MR/K001744/1/MRC_/Medical Research Council/United Kingdom
- BB/J004243/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
