

Regulation of coronavirus nsp15 cleavage specificity by RNA structure
- PMID: 37616296
- PMCID: PMC10449227
- DOI: 10.1371/journal.pone.0290675
Regulation of coronavirus nsp15 cleavage specificity by RNA structure
Abstract
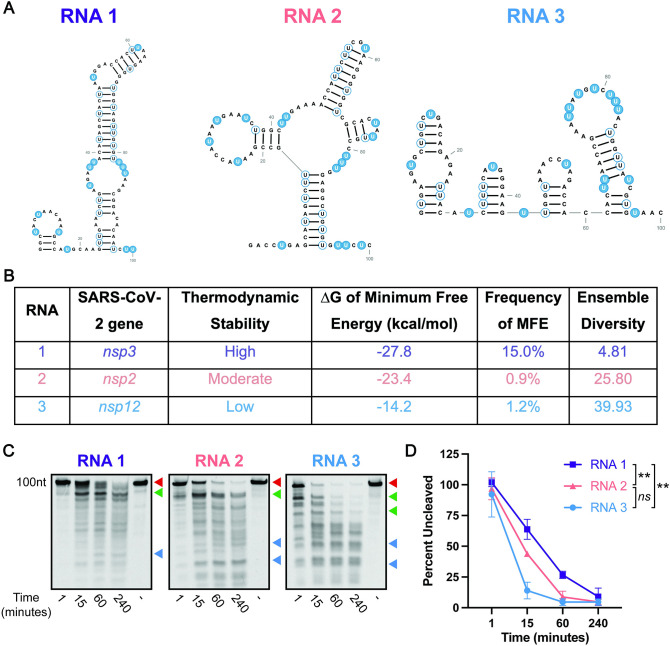
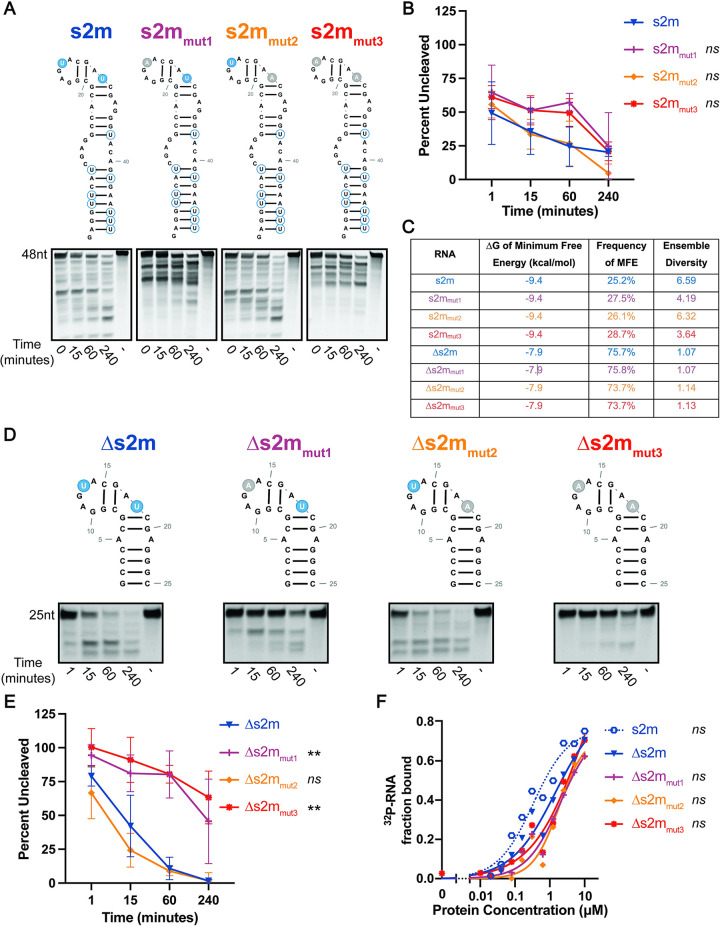
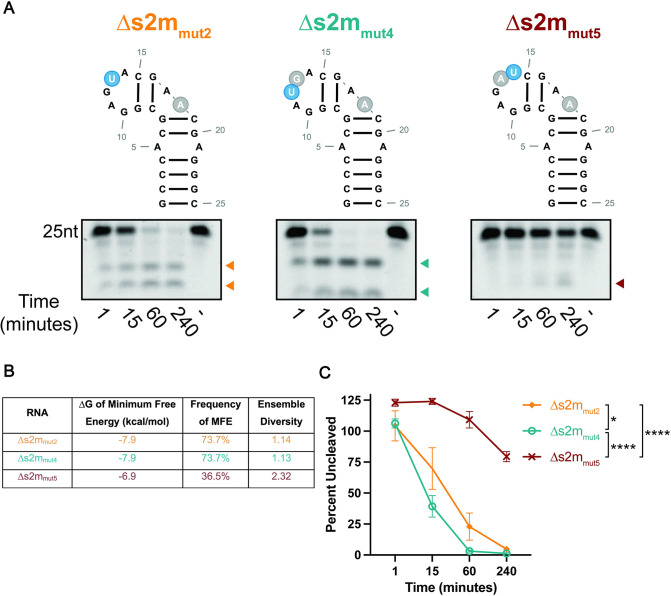
SARS-CoV-2, the etiologic agent of the COVID-19 pandemic, has had an enduring impact on global public health. However, SARS-CoV-2 is only one of multiple pathogenic human coronaviruses (CoVs) to have emerged since the turn of the century. CoVs encode for several nonstructural proteins (nsps) that are essential for viral replication and pathogenesis. Among them is nsp15, a uridine-specific viral endonuclease that is important in evading the host immune response and promoting viral replication. Despite the established endonuclease function of nsp15, little is known about other determinants of its cleavage specificity. In this study we investigate the role of RNA secondary structure in SARS-CoV-2 nsp15 endonuclease activity. Using a series of in vitro endonuclease assays, we observed that thermodynamically stable RNA structures were protected from nsp15 cleavage relative to RNAs lacking stable structure. We leveraged the s2m RNA from the SARS-CoV-1 3'UTR as a model for our structural studies as it adopts a well-defined structure with several uridines, two of which are unpaired and thus highly probable targets for nsp15 cleavage. We found that SARS-CoV-2 nsp15 specifically cleaves s2m at the unpaired uridine within the GNRNA pentaloop of the RNA. Further investigation revealed that the position of uridine within the pentaloop also impacted nsp15 cleavage efficiency suggesting that positioning within the pentaloop is necessary for optimal presentation of the scissile uridine and alignment within the nsp15 catalytic pocket. Our findings indicate that RNA secondary structure is an important determinant of nsp15 cleavage and provides insight into the molecular mechanisms of RNA recognition by nsp15.
Copyright: © 2023 Salukhe et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Helmy YA, Fawzy M, Elaswad A, Sobieh A, Kenney SP, Shehata AA. The COVID-19 Pandemic: A Comprehensive Review of Taxonomy, Genetics, Epidemiology, Diagnosis, Treatment, and Control. J Clin Med. 2020;9(4). Epub 2020/04/30. doi: 10.3390/jcm9041225 ; PubMed Central PMCID: PMC7230578. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
