

The gut microbiota reprograms intestinal lipid metabolism through long noncoding RNA Snhg9
- PMID: 37616368
- PMCID: PMC10688608
- DOI: 10.1126/science.ade0522
The gut microbiota reprograms intestinal lipid metabolism through long noncoding RNA Snhg9
Abstract
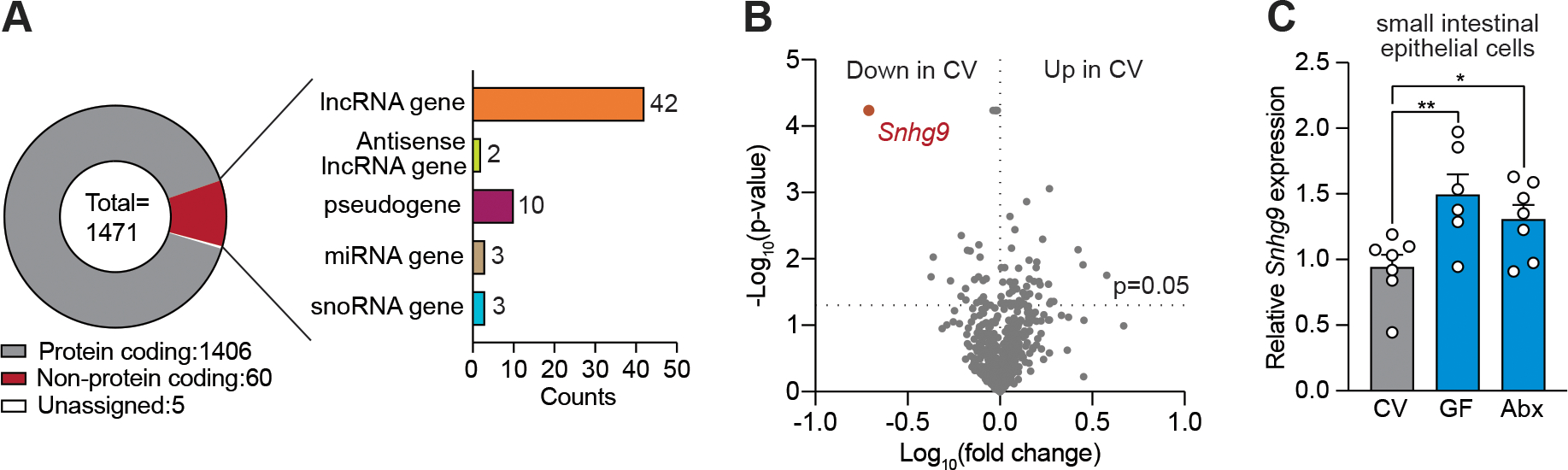
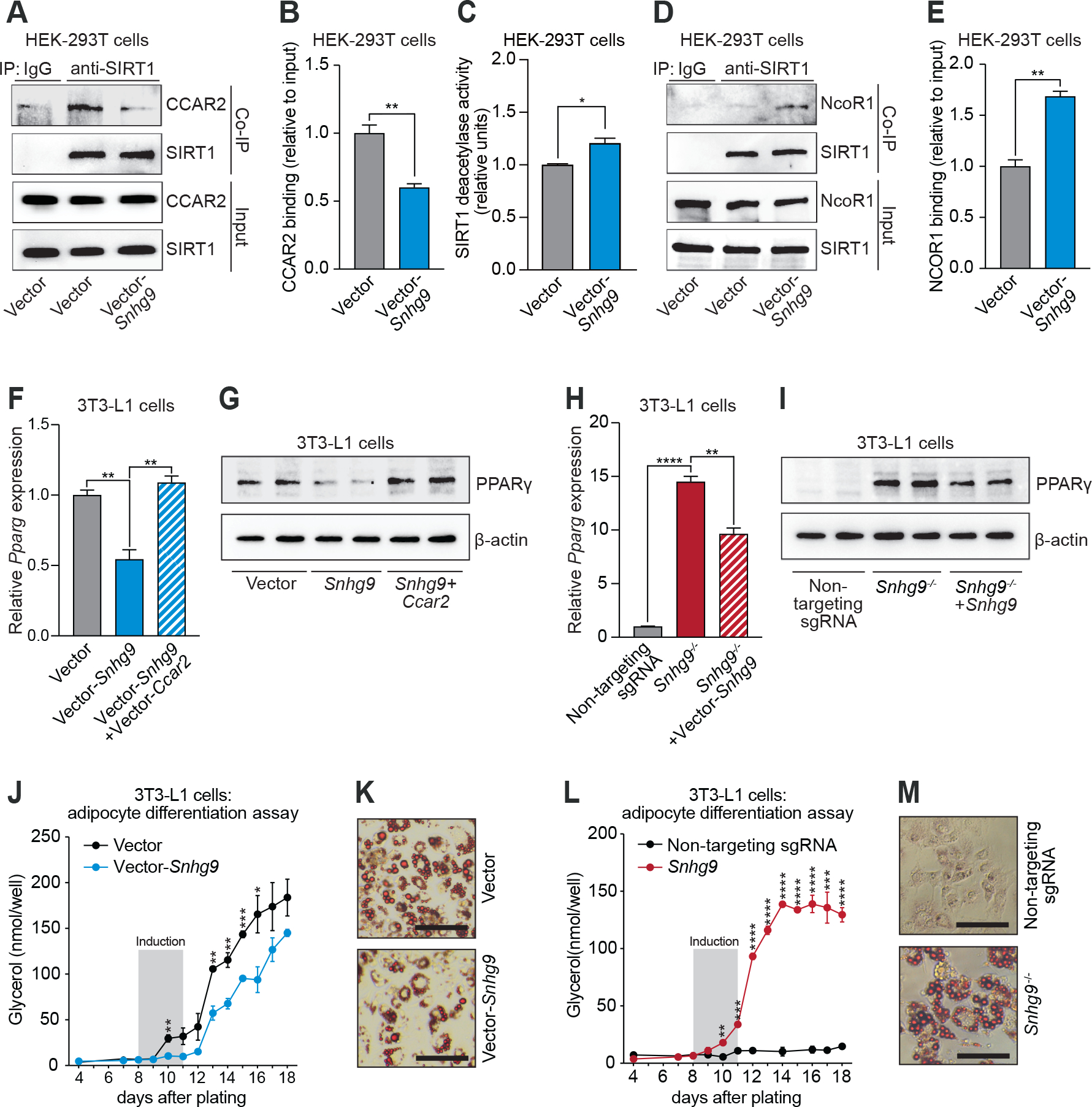
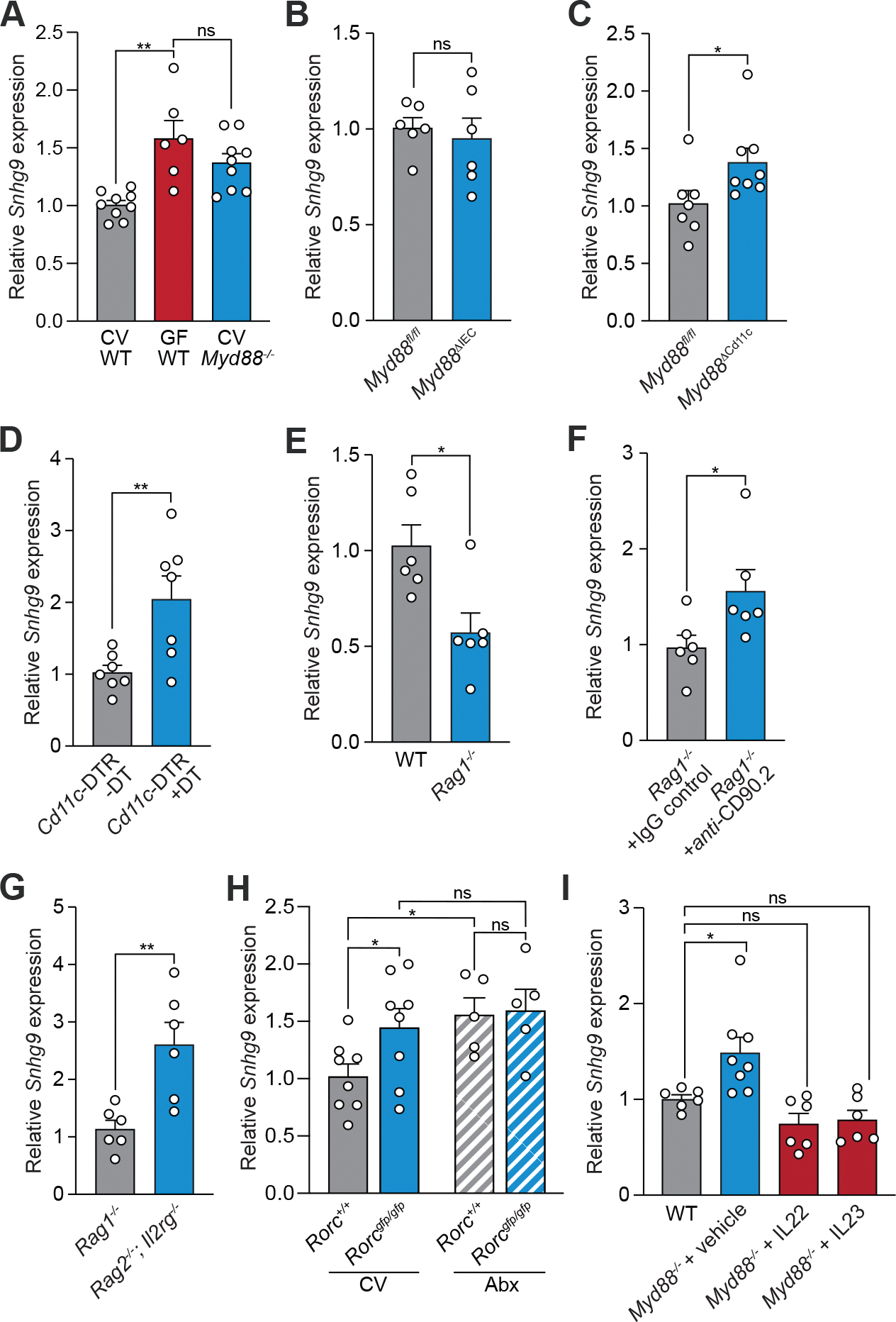
The intestinal microbiota regulates mammalian lipid absorption, metabolism, and storage. We report that the microbiota reprograms intestinal lipid metabolism in mice by repressing the expression of long noncoding RNA (lncRNA) Snhg9 (small nucleolar RNA host gene 9) in small intestinal epithelial cells. Snhg9 suppressed the activity of peroxisome proliferator-activated receptor γ (PPARγ)-a central regulator of lipid metabolism-by dissociating the PPARγ inhibitor sirtuin 1 from cell cycle and apoptosis protein 2 (CCAR2). Forced expression of Snhg9 in the intestinal epithelium of conventional mice impaired lipid absorption, reduced body fat, and protected against diet-induced obesity. The microbiota repressed Snhg9 expression through an immune relay encompassing myeloid cells and group 3 innate lymphoid cells. Our findings thus identify an unanticipated role for a lncRNA in microbial control of host metabolism.
Conflict of interest statement
Figures


References
-
- Hooper LV, Midtvedt T, Gordon JI, How host-microbial interactions shape the nutrient environment of the mammalian intestine. Annu. Rev. Nutr 22, 283–307 (2002). - PubMed
-
- Krautkramer KA, Fan J, Bäckhed F, Gut microbial metabolites as multi-kingdom intermediates. Nat. Rev. Micro. 19, 77–94 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
