

The Architecture of Metabolic Networks Constrains the Evolution of Microbial Resource Hierarchies
- PMID: 37619982
- PMCID: PMC10476156
- DOI: 10.1093/molbev/msad187
The Architecture of Metabolic Networks Constrains the Evolution of Microbial Resource Hierarchies
Abstract
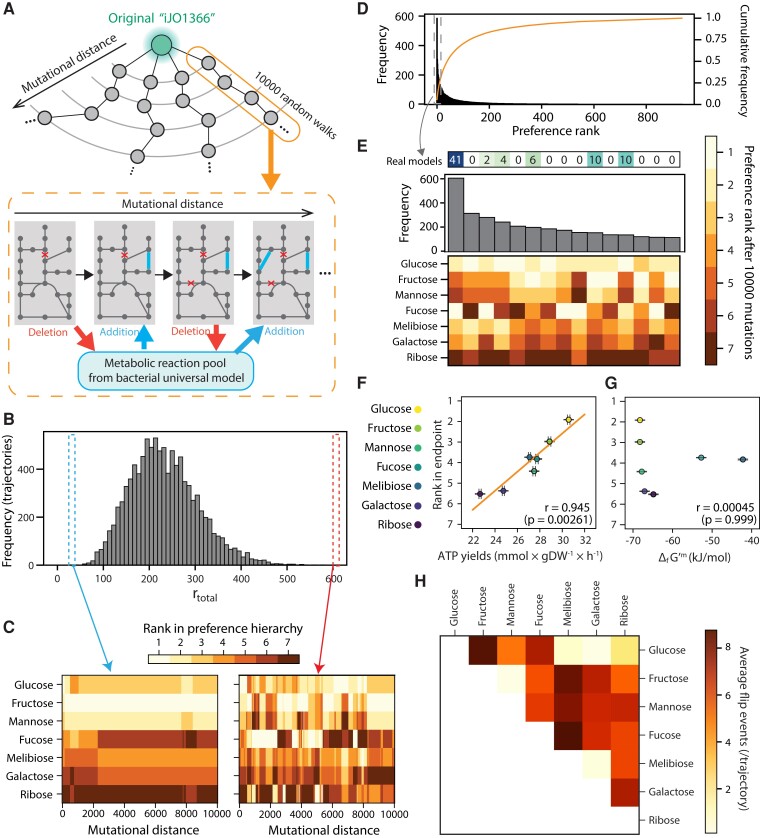
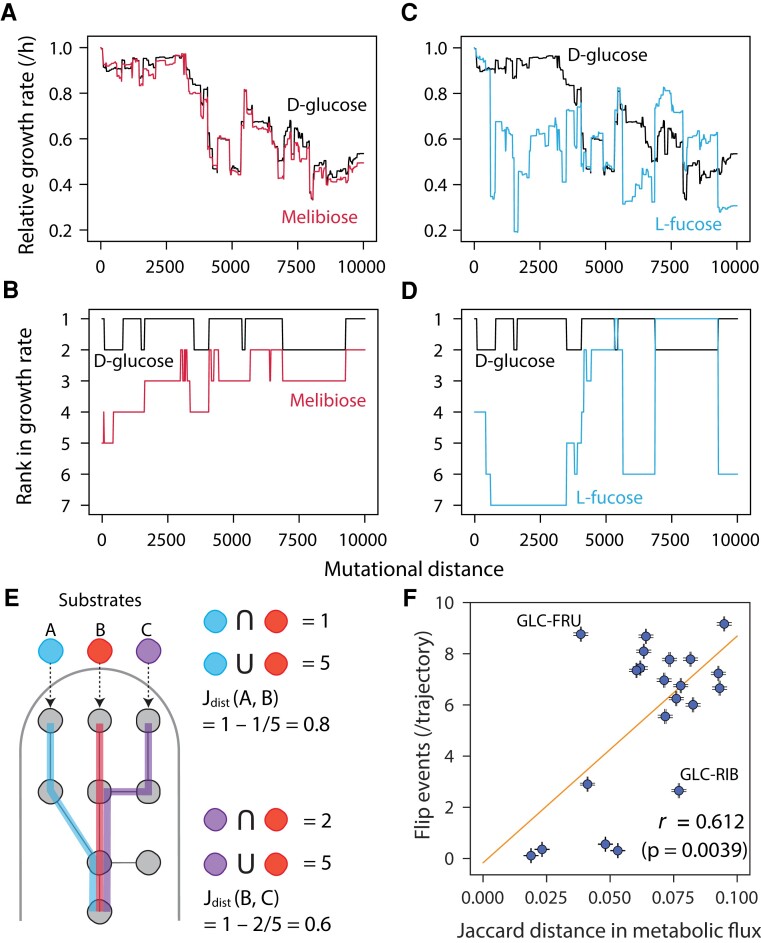
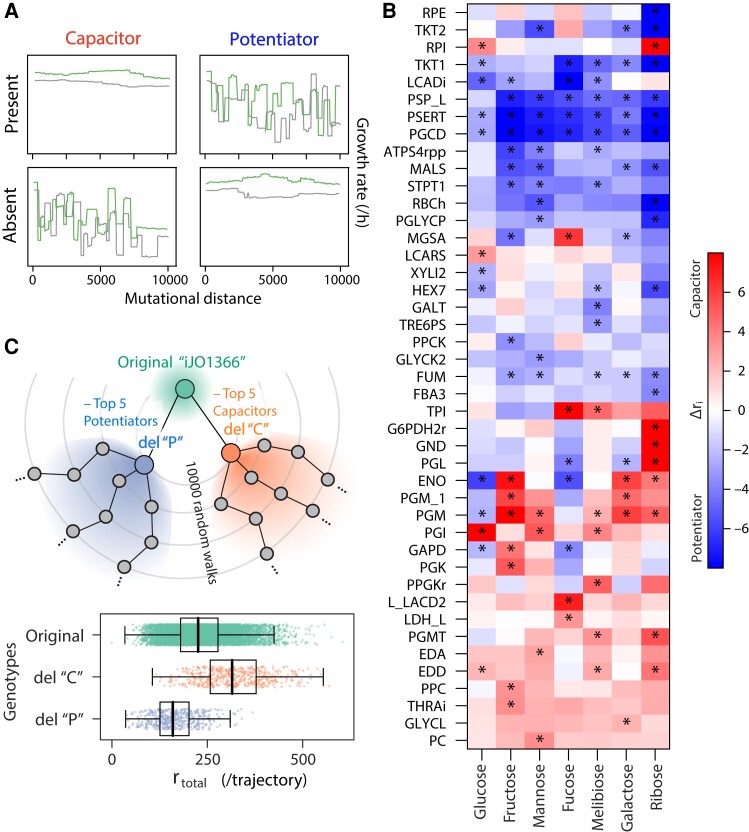
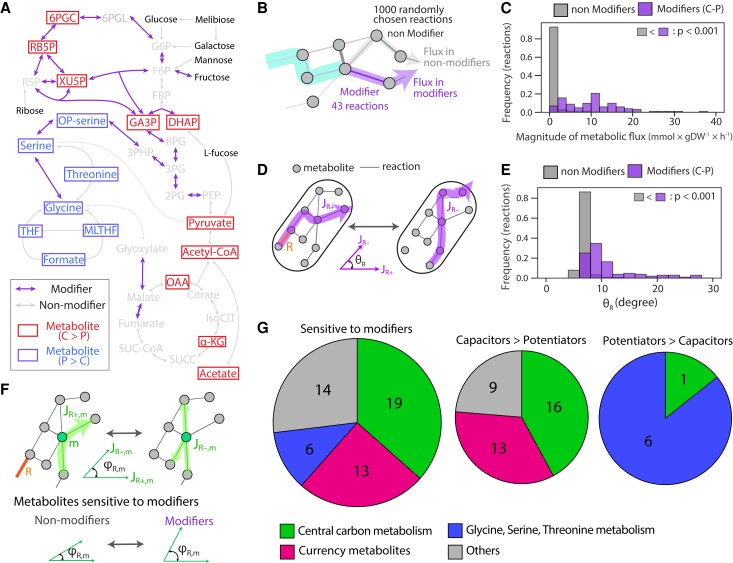
Microbial strategies for resource use are an essential determinant of their fitness in complex habitats. When facing environments with multiple nutrients, microbes often use them sequentially according to a preference hierarchy, resulting in well-known patterns of diauxic growth. In theory, the evolutionary diversification of metabolic hierarchies could represent a mechanism supporting coexistence and biodiversity by enabling temporal segregation of niches. Despite this ecologically critical role, the extent to which substrate preference hierarchies can evolve and diversify remains largely unexplored. Here, we used genome-scale metabolic modeling to systematically explore the evolution of metabolic hierarchies across a vast space of metabolic network genotypes. We find that only a limited number of metabolic hierarchies can readily evolve, corresponding to the most commonly observed hierarchies in genome-derived models. We further show how the evolution of novel hierarchies is constrained by the architecture of central metabolism, which determines both the propensity to change ranks between pairs of substrates and the effect of specific reactions on hierarchy evolution. Our analysis sheds light on the genetic and mechanistic determinants of microbial metabolic hierarchies, opening new research avenues to understand their evolution, evolvability, and ecology.
Keywords: genotype–phenotype maps; metabolic hierarchies; microbial evolution.
© The Author(s) 2023. Published by Oxford University Press on behalf of Society for Molecular Biology and Evolution.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
