

Outer membrane vesicles from a mosquito commensal mediate targeted killing of Plasmodium parasites via the phosphatidylcholine scavenging pathway
- PMID: 37620328
- PMCID: PMC10449815
- DOI: 10.1038/s41467-023-40887-6
Outer membrane vesicles from a mosquito commensal mediate targeted killing of Plasmodium parasites via the phosphatidylcholine scavenging pathway
Abstract
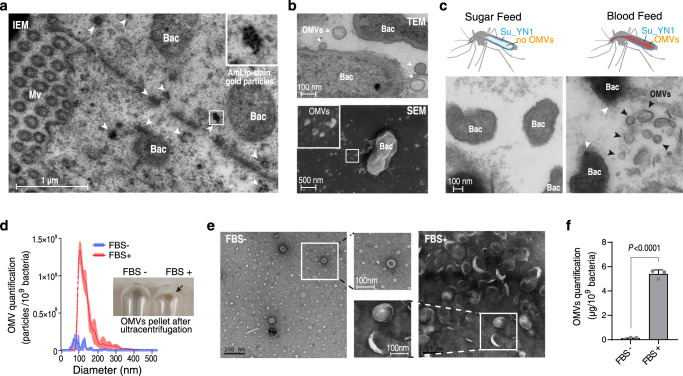
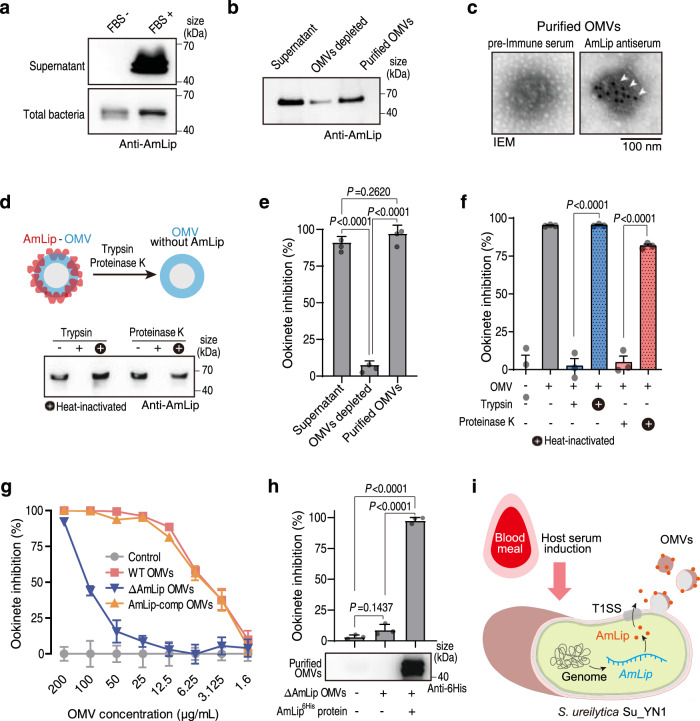
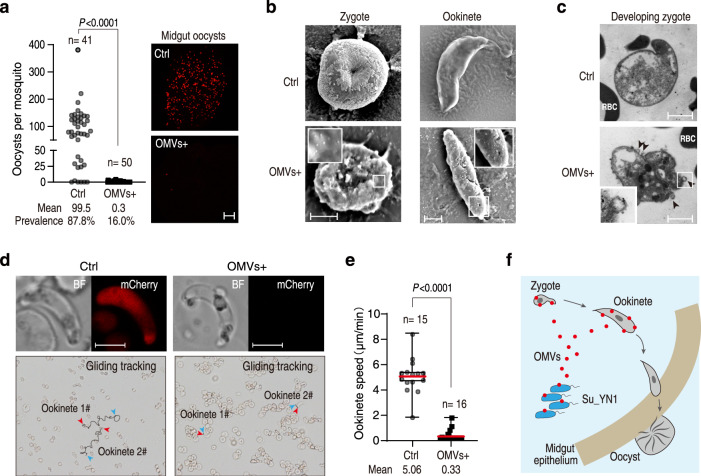
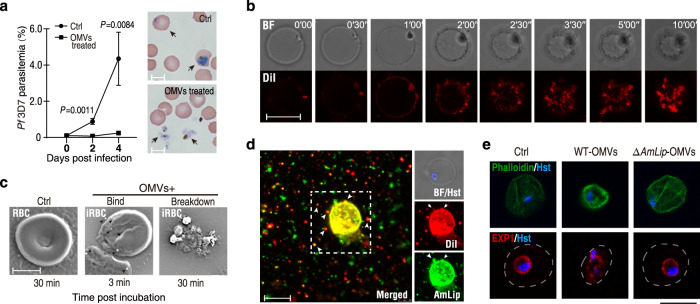
The gut microbiota is a crucial modulator of Plasmodium infection in mosquitoes, including the production of anti-Plasmodium effector proteins. But how the commensal-derived effectors are translocated into Plasmodium parasites remains obscure. Here we show that a natural Plasmodium blocking symbiotic bacterium Serratia ureilytica Su_YN1 delivers the effector lipase AmLip to Plasmodium parasites via outer membrane vesicles (OMVs). After a blood meal, host serum strongly induces Su_YN1 to release OMVs and the antimalarial effector protein AmLip into the mosquito gut. AmLip is first secreted into the extracellular space via the T1SS and then preferentially loaded on the OMVs that selectively target the malaria parasite, leading to targeted killing of the parasites. Notably, these serum-induced OMVs incorporate certain serum-derived lipids, such as phosphatidylcholine, which is critical for OMV uptake by Plasmodium via the phosphatidylcholine scavenging pathway. These findings reveal that this gut symbiotic bacterium evolved to deliver secreted effector molecules in the form of extracellular vesicles to selectively attack parasites and render mosquitoes refractory to Plasmodium infection. The discovery of the role of gut commensal-derived OMVs as carriers in cross-kingdom communication between mosquito microbiota and Plasmodium parasites offers a potential innovative strategy for blocking malaria transmission.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
A natural symbiotic bacterium drives mosquito refractoriness to Plasmodium infection via secretion of an antimalarial lipase.Nat Microbiol. 2021 Jun;6(6):806-817. doi: 10.1038/s41564-021-00899-8. Epub 2021 May 6. Nat Microbiol. 2021. PMID: 33958765 Free PMC article.
-
Quorum sensing-activated phenylalanine metabolism drives OMV biogenesis to enhance mosquito commensal colonization resistance to Plasmodium.Cell Host Microbe. 2023 Oct 11;31(10):1655-1667.e6. doi: 10.1016/j.chom.2023.08.017. Epub 2023 Sep 21. Cell Host Microbe. 2023. PMID: 37738984
-
Characterization and proteomic analysis of outer membrane vesicles from a commensal microbe, Enterobacter cloacae.J Proteomics. 2021 Jan 16;231:103994. doi: 10.1016/j.jprot.2020.103994. Epub 2020 Sep 29. J Proteomics. 2021. PMID: 33007464
-
Transmission-Blocking Strategies Against Malaria Parasites During Their Mosquito Stages.Front Cell Infect Microbiol. 2022 Feb 16;12:820650. doi: 10.3389/fcimb.2022.820650. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35252033 Free PMC article. Review.
-
The microbiota, the malarial parasite, and the mosquito [MMM] - A three-sided relationship.Mol Biochem Parasitol. 2023 Feb;253:111543. doi: 10.1016/j.molbiopara.2023.111543. Epub 2023 Jan 13. Mol Biochem Parasitol. 2023. PMID: 36642385 Review.
Cited by
-
Bacterial membrane vesicles: orchestrators of interkingdom interactions in microbial communities for environmental adaptation and pathogenic dynamics.Front Immunol. 2024 Mar 21;15:1371317. doi: 10.3389/fimmu.2024.1371317. eCollection 2024. Front Immunol. 2024. PMID: 38576623 Free PMC article. Review.
-
Algicidal bacteria-derived membrane vesicles as shuttles mediating cross-kingdom interactions between bacteria and algae.Sci Adv. 2024 Aug 9;10(32):eadn4526. doi: 10.1126/sciadv.adn4526. Epub 2024 Aug 7. Sci Adv. 2024. PMID: 39110793 Free PMC article.
References
-
- Roberts L. Lingering fever. Science. 2022;377:1139–1143. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
