

A synthetic cell-free 36-enzyme reaction system for vitamin B12 production
- PMID: 37620358
- PMCID: PMC10449867
- DOI: 10.1038/s41467-023-40932-4
A synthetic cell-free 36-enzyme reaction system for vitamin B12 production
Abstract
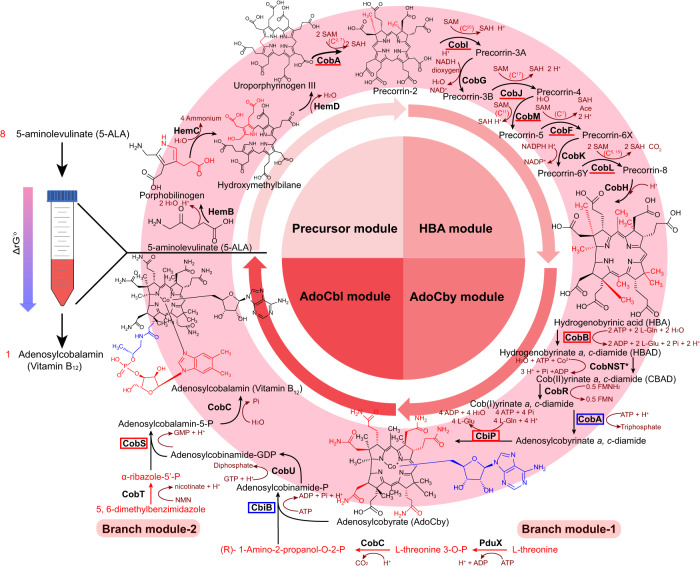
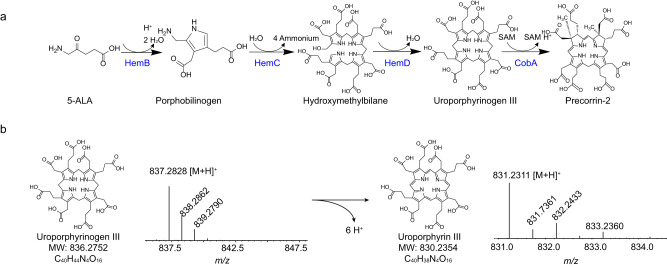
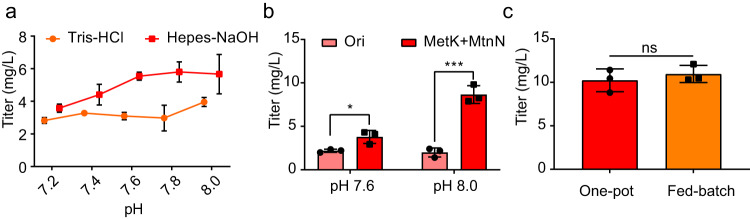
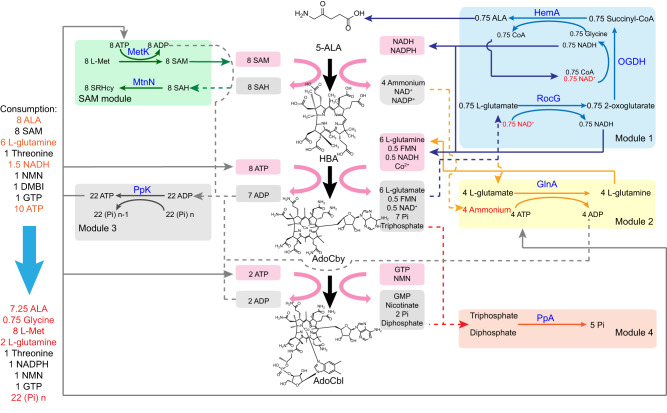
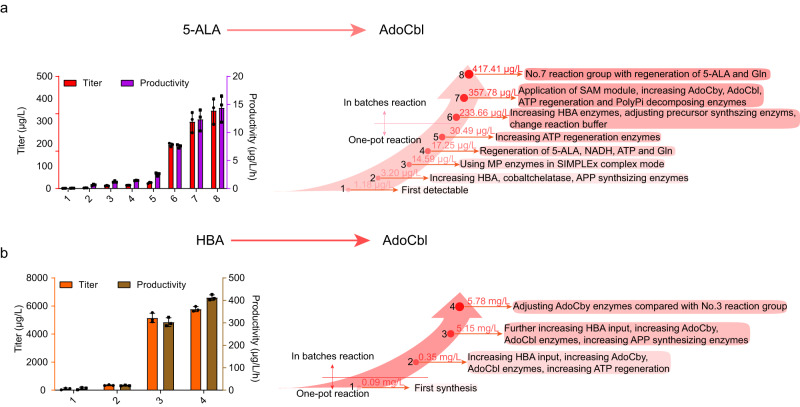
Adenosylcobalamin (AdoCbl), a biologically active form of vitamin B12 (coenzyme B12), is one of the most complex metal-containing natural compounds and an essential vitamin for animals. However, AdoCbl can only be de novo synthesized by prokaryotes, and its industrial manufacturing to date was limited to bacterial fermentation. Here, we report a method for the synthesis of AdoCbl based on a cell-free reaction system performing a cascade of catalytic reactions from 5-aminolevulinic acid (5-ALA), an inexpensive compound. More than 30 biocatalytic reactions are integrated and optimized to achieve the complete cell-free synthesis of AdoCbl, after overcoming feedback inhibition, the complicated detection, instability of intermediate products, as well as imbalance and competition of cofactors. In the end, this cell-free system produces 417.41 μg/L and 5.78 mg/L of AdoCbl using 5-ALA and the purified intermediate product hydrogenobyrate as substrates, respectively. The strategies of coordinating synthetic modules of complex cell-free system describe here will be generally useful for developing cell-free platforms to produce complex natural compounds with long and complicated biosynthetic pathways.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Green R, et al. Vitamin B(12) deficiency. Nat. Rev. Dis. Prim. 2017;3:17040. - PubMed
-
- Woodward RB. The total synthesis of vitamin B 12. Pure Appl. Chem. 1973;33:145–177. - PubMed
-
- Wang ZJ, et al. Improved vitamin B(12) production by step-wise reduction of oxygen uptake rate under dissolved oxygen limiting level during fermentation process. Bioresor Technol. 2010;101:2845–2852. - PubMed
-
- Wang P, Zhang Z, Jiao Y, Liu S, Wang Y. Improved propionic acid and 5,6-dimethylbenzimidazole control strategy for vitamin B12 fermentation by Propionibacterium freudenreichii. J. Biotechnol. 2015;193:123–129. - PubMed
-
- Dong H, et al. A newly isolated and identified vitamin B12 producing strain: Sinorhizobium meliloti 320. Bioprocess Biosyst. Eng. 2016;39:1527–1537. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
