

High-precision mapping reveals the structure of odor coding in the human brain
- PMID: 37620443
- PMCID: PMC10726579
- DOI: 10.1038/s41593-023-01414-4
High-precision mapping reveals the structure of odor coding in the human brain
Abstract
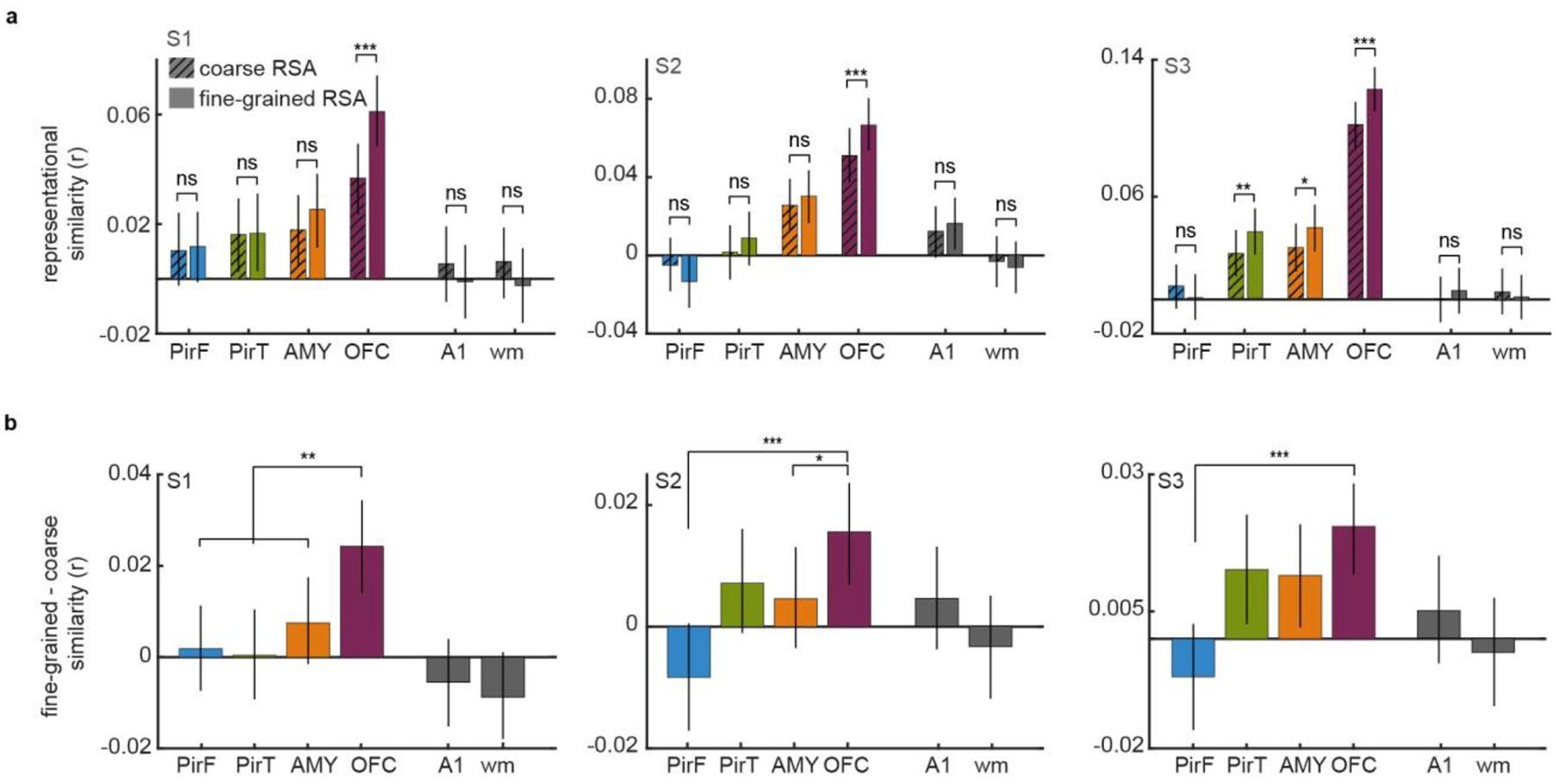
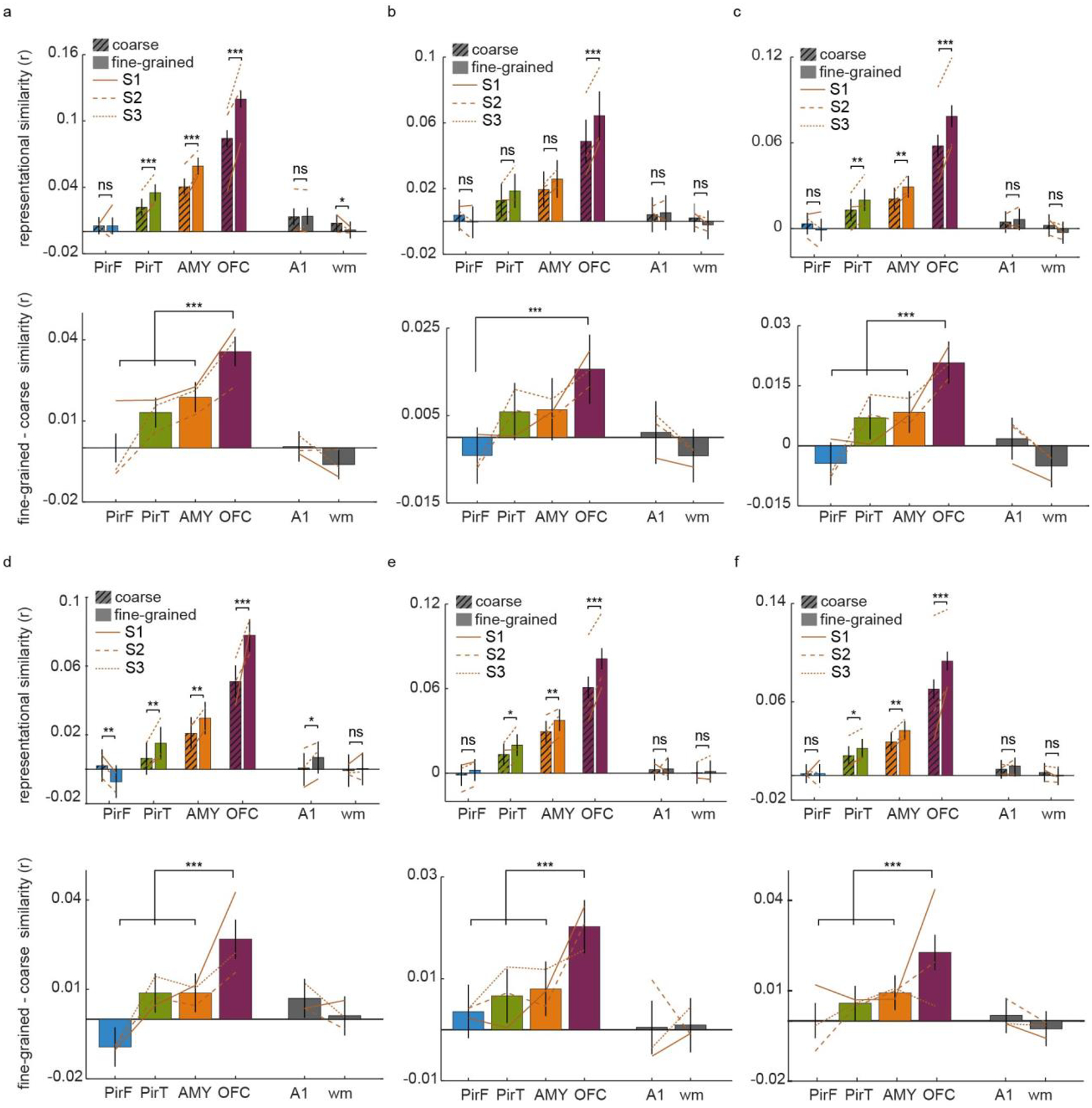
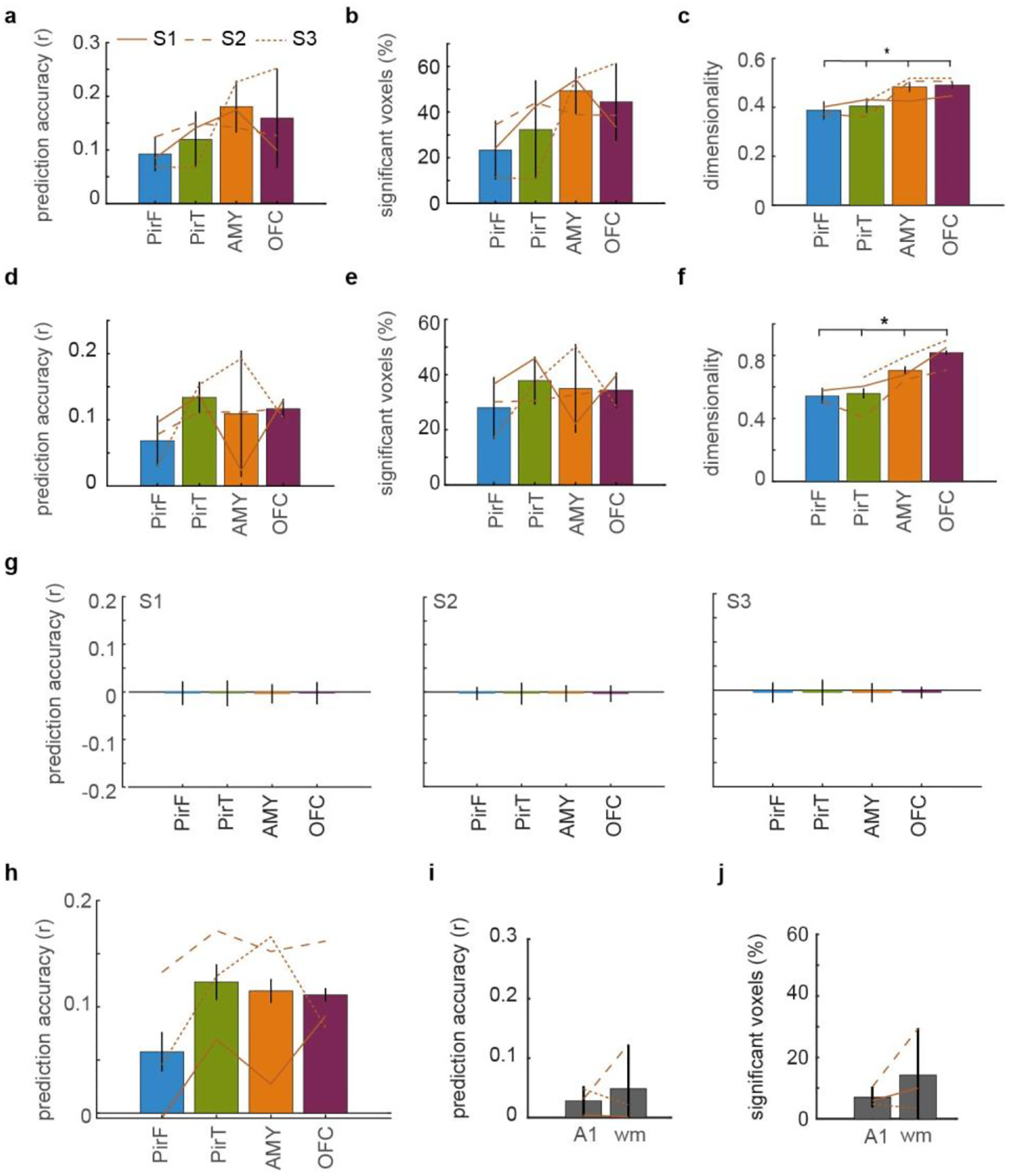
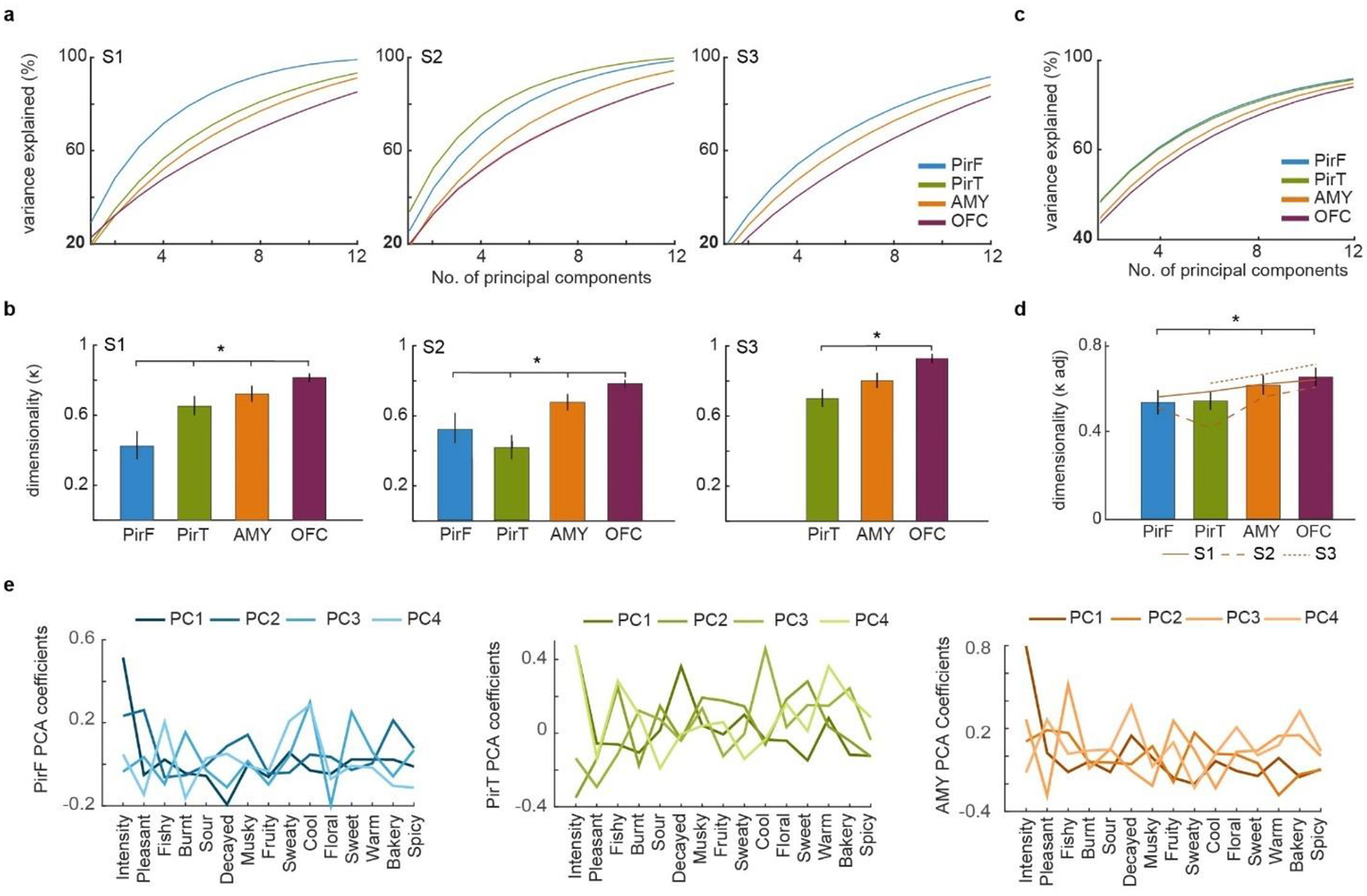
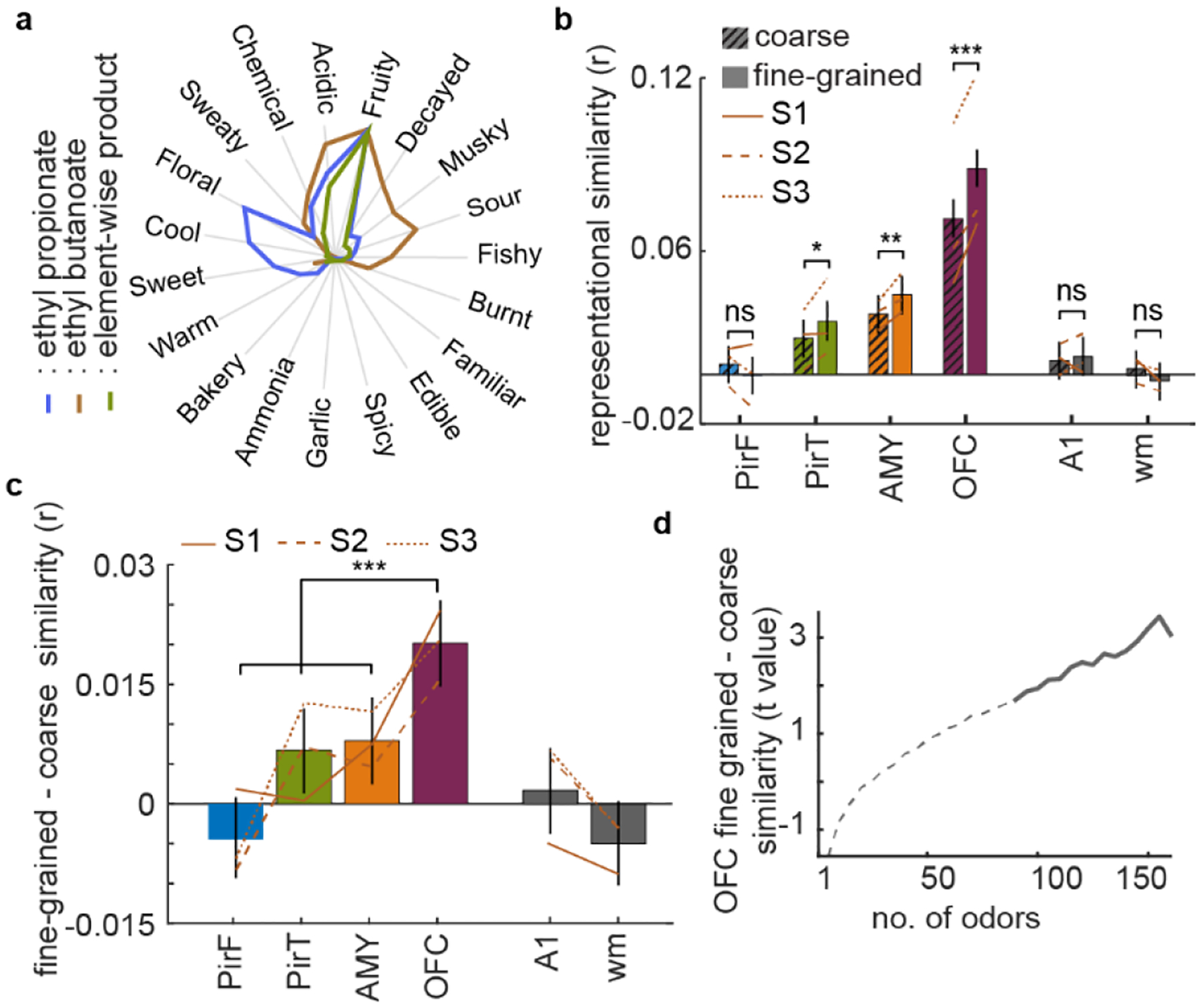
Odor perception is inherently subjective. Previous work has shown that odorous molecules evoke distributed activity patterns in olfactory cortices, but how these patterns map on to subjective odor percepts remains unclear. In the present study, we collected neuroimaging responses to 160 odors from 3 individual subjects (18 h per subject) to probe the neural coding scheme underlying idiosyncratic odor perception. We found that activity in the orbitofrontal cortex (OFC) represents the fine-grained perceptual identity of odors over and above coarsely defined percepts, whereas this difference is less pronounced in the piriform cortex (PirC) and amygdala. Furthermore, the implementation of perceptual encoding models enabled us to predict olfactory functional magnetic resonance imaging responses to new odors, revealing that the dimensionality of the encoded perceptual spaces increases from the PirC to the OFC. Whereas encoding of lower-order dimensions generalizes across subjects, encoding of higher-order dimensions is idiosyncratic. These results provide new insights into cortical mechanisms of odor coding and suggest that subjective olfactory percepts reside in the OFC.
© 2023. This is a U.S. Government work and not under copyright protection in the US; foreign copyright protection may apply.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures










References
-
- Leopold DA, Wilke M, Maier A & Logothetis NK Stable perception of visually ambiguous patterns. Nature neuroscience 5, 605–609 (2002). - PubMed
-
- Walsh V & Kulikowski J Perceptual constancy: Why things look as they do. (Cambridge University Press, 1998).
-
- Hudson R From molecule to mind: the role of experience in shaping olfactory function. Journal of Comparative Physiology A 185, 297–304 (1999). - PubMed
-
- Sell CS On the unpredictability of odor. Angewandte Chemie International Edition 45, 6254–6261 (2006). - PubMed
METHOD REFERENCES
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
