

The Extracellular Lipopeptides and Volatile Organic Compounds of Bacillus subtilis DHA41 Display Broad-Spectrum Antifungal Activity against Soil-Borne Phytopathogenic Fungi
- PMID: 37623568
- PMCID: PMC10455929
- DOI: 10.3390/jof9080797
The Extracellular Lipopeptides and Volatile Organic Compounds of Bacillus subtilis DHA41 Display Broad-Spectrum Antifungal Activity against Soil-Borne Phytopathogenic Fungi
Abstract
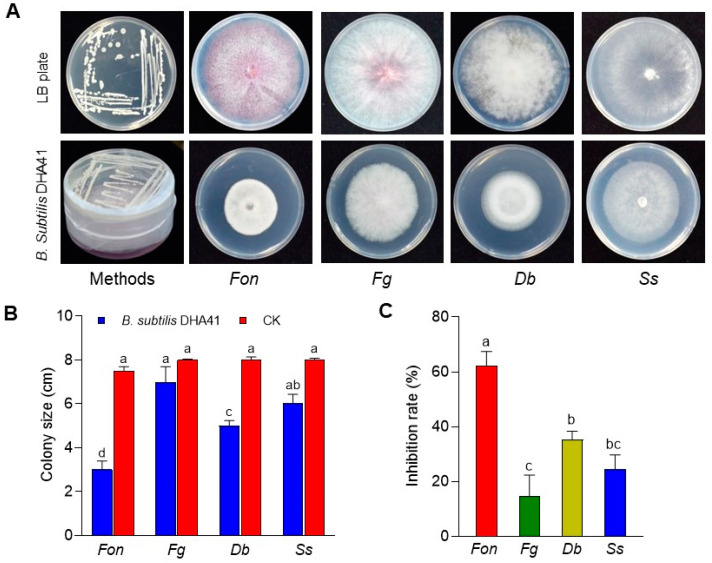
Fusarium oxysporum f. sp. niveum (Fon) is a devastating soil-borne fungus causing Fusarium wilt in watermelon. The present study investigated the biochemical mechanism underlying the antifungal activity exhibited by the antagonistic bacterial strain DHA41, particularly against Fon. Molecular characterization based on the 16S rRNA gene confirmed that DHA41 is a strain of Bacillus subtilis, capable of synthesizing antifungal lipopeptides, such as iturins and fengycins, which was further confirmed by detecting corresponding lipopeptide biosynthesis genes, namely ItuB, ItuD, and FenD. The cell-free culture filtrate and extracellular lipopeptide extract of B. subtilis DHA41 demonstrated significant inhibitory effects on the mycelial growth of Fon, Didymella bryoniae, Sclerotinia sclerotiorum, Fusarium graminearum, and Rhizoctonia solani. The lipopeptide extract showed emulsification activity and inhibited Fon mycelial growth by 86.4% at 100 µg/mL. Transmission electron microscope observations confirmed that the lipopeptide extract disrupted Fon cellular integrity. Furthermore, B. subtilis DHA41 emitted volatile organic compounds (VOCs) that exhibited antifungal activity against Fon, D. bryoniae, S. sclerotiorum, and F. graminearum. These findings provide evidence that B. subtilis DHA41 possesses broad-spectrum antifungal activity against different fungi pathogens, including Fon, through the production of extracellular lipopeptides and VOCs.
Keywords: Fusarium oxysporum f. sp. niveum; antifungal activity; lipopeptide; soil-borne pathogens; volatile organic compounds.
Conflict of interest statement
The authors declare no conflict of interest.
Figures







References
-
- Coque J.J.R., Álvarez-Pérez J.M., Cobos R., González-García S., Ibáñez A.M., Diez Galán A., Calvo-Peña C. Advances in the control of phytopathogenic fungi that infect crops through their root system. Adv. Appl. Microbiol. 2020;111:123–170. - PubMed
-
- Lü G., Guo S., Zhang H., Geng L., Song F., Fei Z., Xu Y. Transcriptional profiling of watermelon during its incompatible interaction with Fusarium oxysporum f. sp. niveum. Eur. J. Plant Pathol. 2011;131:585–601. doi: 10.1007/s10658-011-9833-z. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
