

Identification of a common secondary mutation in the Neurospora crassa knockout collection conferring a cell fusion-defective phenotype
- PMID: 37623742
- PMCID: PMC10580951
- DOI: 10.1128/spectrum.02087-23
Identification of a common secondary mutation in the Neurospora crassa knockout collection conferring a cell fusion-defective phenotype
Abstract
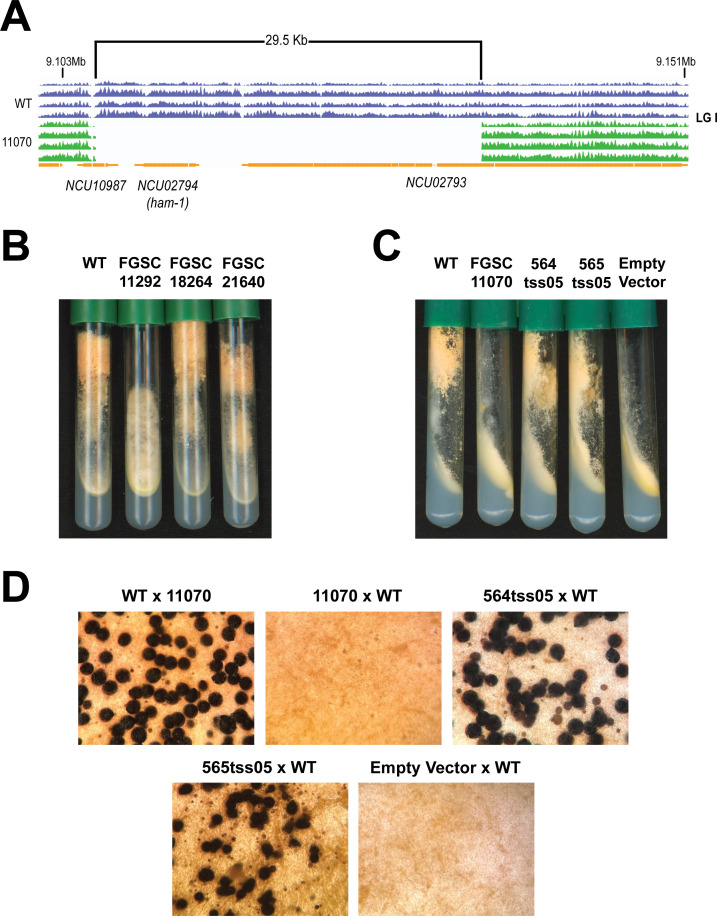
Gene-deletion mutants represent a powerful tool to study gene function. The filamentous fungus Neurospora crassa is a well-established model organism, and features a comprehensive gene knockout strain collection. While these mutant strains have been used in numerous studies, resulting in the functional annotation of many Neurospora genes, direct confirmation of gene-phenotype relationships is often lacking, which is particularly relevant given the possibility of background mutations, sample contamination, and/or strain mislabeling. Indeed, spontaneous mutations resulting in phenotypes resembling many cell fusion mutants have long been known to occur at relatively high frequency in N. crassa, and these secondary mutations are common in the Neurospora deletion collection. The identity of these mutations, however, is largely unknown. Here, we report that the Δada-3 strain from the N. crassa knockout collection, which exhibits a cell fusion defect, harbors a secondary mutation responsible for this phenotype. Through whole-genome sequencing and genetic analyses, we found a ~30-Kb deletion in this strain affecting a known cell fusion-related gene, so/ham-1, and show that it is the absence of this gene-and not of ada-3-that underlies its cell fusion defect. We additionally found three other knockout strains harboring the same deletion, suggesting that this mutation may be common in the collection and could have impacted previous studies. Our findings provide a cautionary note and highlight the importance of proper functional validation of strains from mutant collections. We discuss our results in the context of the spread of cell fusion-defective cheater variants in N. crassa cultures. IMPORTANCE This study emphasizes the need for careful and detailed characterization of strains from mutant collections. Specifically, we found a common deletion in various strains from the Neurospora crassa gene knockout collection that results in a cell fusion-defective phenotype. This is noteworthy because this collection is known to contain background mutations-of a largely unclear nature-that produce cell fusion-defective phenotypes. Our results describe an example of such mutations, and highlight how this common genetic defect could have impacted previous studies that have used the affected strains. Furthermore, they provide a cautionary note about the use of Neurospora strains with similar phenotypes. Lastly, these findings offer additional details relevant to our understanding of the origin and spread of cell fusion-defective cheater variants in N. crassa cultures.
Keywords: Neurospora crassa; cell fusion; gene knockout; secondary mutations.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



Similar articles
-
Clustering analysis of large-scale phenotypic data in the model filamentous fungus Neurospora crassa.BMC Genomics. 2020 Nov 2;21(1):755. doi: 10.1186/s12864-020-07131-7. BMC Genomics. 2020. PMID: 33138786 Free PMC article.
-
The Saccharomyces cerevisiae PRM1 homolog in Neurospora crassa is involved in vegetative and sexual cell fusion events but also has postfertilization functions.Genetics. 2009 Feb;181(2):497-510. doi: 10.1534/genetics.108.096149. Epub 2008 Dec 8. Genetics. 2009. PMID: 19064710 Free PMC article.
-
Deletion of homologs of the SREBP pathway results in hyper-production of cellulases in Neurospora crassa and Trichoderma reesei.Biotechnol Biofuels. 2015 Aug 19;8:121. doi: 10.1186/s13068-015-0297-9. eCollection 2015. Biotechnol Biofuels. 2015. PMID: 26288653 Free PMC article.
-
Genetic control of radiation sensitivity and DNA repair in Neurospora.Basic Life Sci. 1975;5B:567-76. doi: 10.1007/978-1-4684-2898-8_22. Basic Life Sci. 1975. PMID: 1103873 Review.
-
Cell fusion in the filamentous fungus, Neurospora crassa.Methods Mol Biol. 2008;475:21-38. doi: 10.1007/978-1-59745-250-2_2. Methods Mol Biol. 2008. PMID: 18979236 Review.
Cited by
-
Considerations for Domestication of Novel Strains of Filamentous Fungi.ACS Synth Biol. 2025 Feb 21;14(2):343-362. doi: 10.1021/acssynbio.4c00672. Epub 2025 Jan 30. ACS Synth Biol. 2025. PMID: 39883596 Free PMC article. Review.
-
Unveiling a classical mutant in the context of the GH3 β-glucosidase family in Neurospora crassa.AMB Express. 2024 Jan 5;14(1):4. doi: 10.1186/s13568-023-01658-0. AMB Express. 2024. PMID: 38180602 Free PMC article.
References
-
- Davis RH. 2000. Neurospora: contributions of a model organism. Oxford University Press.
-
- Galagan JE, Calvo SE, Borkovich KA, Selker EU, Read ND, Jaffe D, FitzHugh W, Ma L-J, Smirnov S, Purcell S, Rehman B, Elkins T, Engels R, Wang S, Nielsen CB, Butler J, Endrizzi M, Qui D, Ianakiev P, Bell-Pedersen D, Nelson MA, Werner-Washburne M, Selitrennikoff CP, Kinsey JA, Braun EL, Zelter A, Schulte U, Kothe GO, Jedd G, Mewes W, Staben C, Marcotte E, Greenberg D, Roy A, Foley K, Naylor J, Stange-Thomann N, Barrett R, Gnerre S, Kamal M, Kamvysselis M, Mauceli E, Bielke C, Rudd S, Frishman D, Krystofova S, Rasmussen C, Metzenberg RL, Perkins DD, Kroken S, Cogoni C, Macino G, Catcheside D, Li W, Pratt RJ, Osmani SA, DeSouza CPC, Glass L, Orbach MJ, Berglund JA, Voelker R, Yarden O, Plamann M, Seiler S, Dunlap J, Radford A, Aramayo R, Natvig DO, Alex LA, Mannhaupt G, Ebbole DJ, Freitag M, Paulsen I, Sachs MS, Lander ES, Nusbaum C, Birren B. 2003. The genome sequence of the filamentous fungus Neurospora crassa. Nature 422:859–868. doi: 10.1038/nature01554 - DOI - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
