

Transcriptional regulation by the NSL complex enables diversification of IFT functions in ciliated versus nonciliated cells
- PMID: 37624894
- PMCID: PMC10456878
- DOI: 10.1126/sciadv.adh5598
Transcriptional regulation by the NSL complex enables diversification of IFT functions in ciliated versus nonciliated cells
Abstract
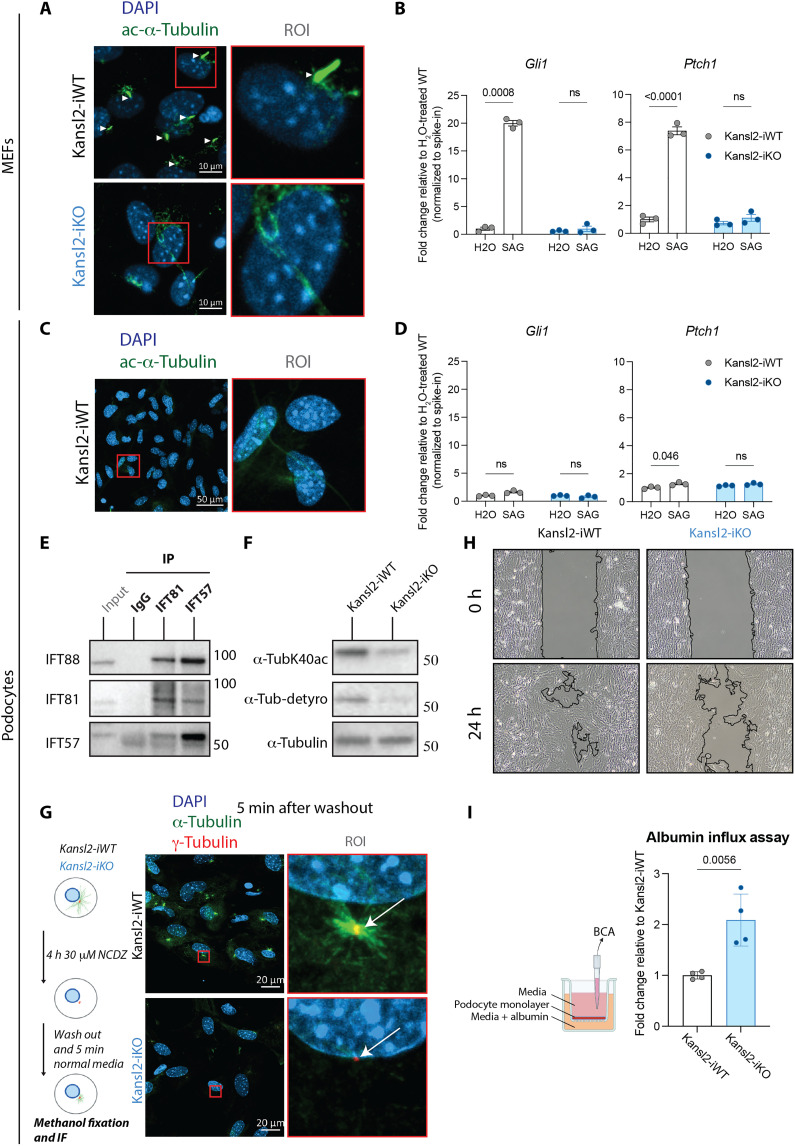
Members of the NSL histone acetyltransferase complex are involved in multiorgan developmental syndromes. While the NSL complex is known for its importance in early development, its role in fully differentiated cells remains enigmatic. Using a kidney-specific model, we discovered that deletion of NSL complex members KANSL2 or KANSL3 in postmitotic podocytes led to catastrophic kidney dysfunction. Systematic comparison of two primary differentiated cell types reveals the NSL complex as a master regulator of intraciliary transport genes in both dividing and nondividing cells. NSL complex ablation led to loss of cilia and impaired sonic hedgehog pathway in ciliated fibroblasts. By contrast, nonciliated podocytes responded with altered microtubule dynamics and obliterated podocyte functions. Finally, overexpression of wild-type but not a double zinc finger (ZF-ZF) domain mutant of KANSL2 rescued the transcriptional defects, revealing a critical function of this domain in NSL complex assembly and function. Thus, the NSL complex exhibits bifurcation of functions to enable diversity of specialized outcomes in differentiated cells.
Figures





References
-
- B. N. Sheikh, A. Akhtar, The many lives of KATs—Detectors, integrators and modulators of the cellular environment. Nat. Rev. Genet. 20, 7–23 (2018). - PubMed
-
- X.-J. Yang, E. Seto, HATs and HDACs: From structure, function and regulation to novel strategies for therapy and prevention. Oncogene 26, 5310–5318 (2007). - PubMed
-
- L. Li, M. Ghorbani, M. Weisz-Hubshman, J. Rousseau, I. Thiffault, R. E. Schnur, C. Breen, R. Oegema, M. M. Weiss, Q. Waisfisz, S. Welner, H. Kingston, J. A. Hills, E. M. Boon, L. Basel-Salmon, O. Konen, H. Goldberg-Stern, L. Bazak, S. Tzur, J. Jin, X. Bi, M. Bruccoleri, K. McWalter, M. T. Cho, M. Scarano, G. B. Schaefer, S. S. Brooks, S. S. Hughes, K. L. I. van Gassen, J. M. van Hagen, T. K. Pandita, P. B. Agrawal, P. M. Campeau, X.-J. Yang, Lysine acetyltransferase 8 is involved in cerebral development and syndromic intellectual disability. J. Clin. Invest. 130, 1431–1445 (2020). - PMC - PubMed
-
- J. H. Roelfsema, S. J. White, Y. Ariyürek, D. Bartholdi, D. Niedrist, F. Papadia, C. A. Bacino, J. T. den Dunnen, G.-J. B. van Ommen, M. H. Breuning, R. C. Hennekam, D. J. M. Peters, Genetic heterogeneity in Rubinstein-Taybi syndrome: Mutations in both the CBP and EP300 genes cause disease. Am. J. Hum. Genet. 76, 572–580 (2005). - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
