

Sustained CD28 costimulation is required for self-renewal and differentiation of TCF-1+ PD-1+ CD8 T cells
- PMID: 37624910
- PMCID: PMC10805182
- DOI: 10.1126/sciimmunol.adg0878
Sustained CD28 costimulation is required for self-renewal and differentiation of TCF-1+ PD-1+ CD8 T cells
Abstract
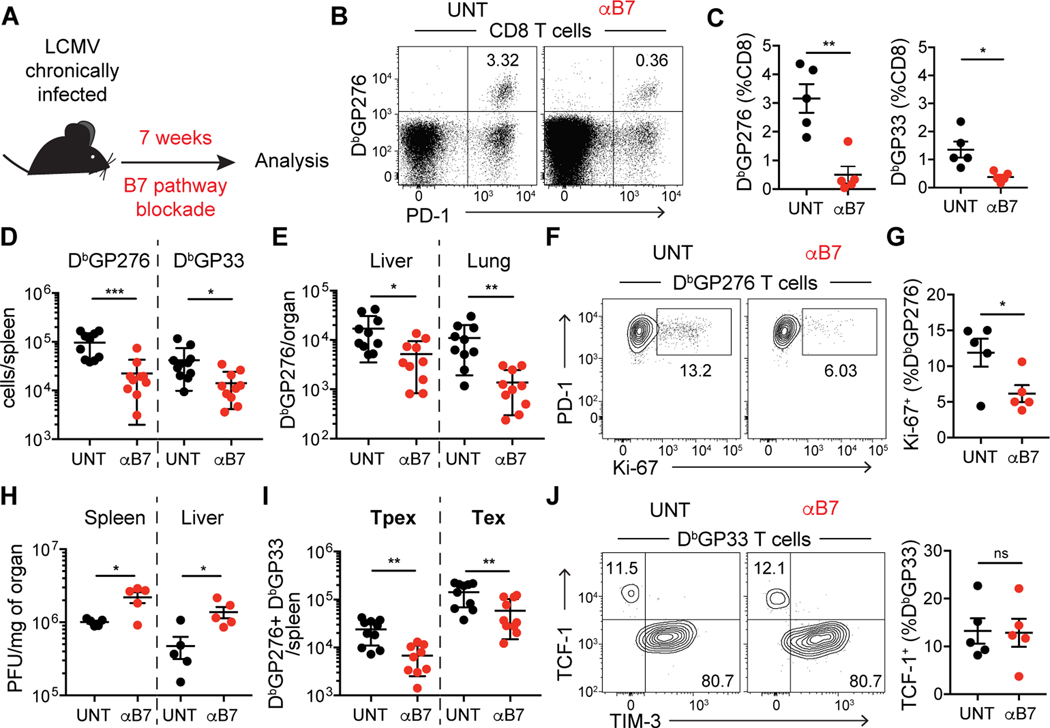
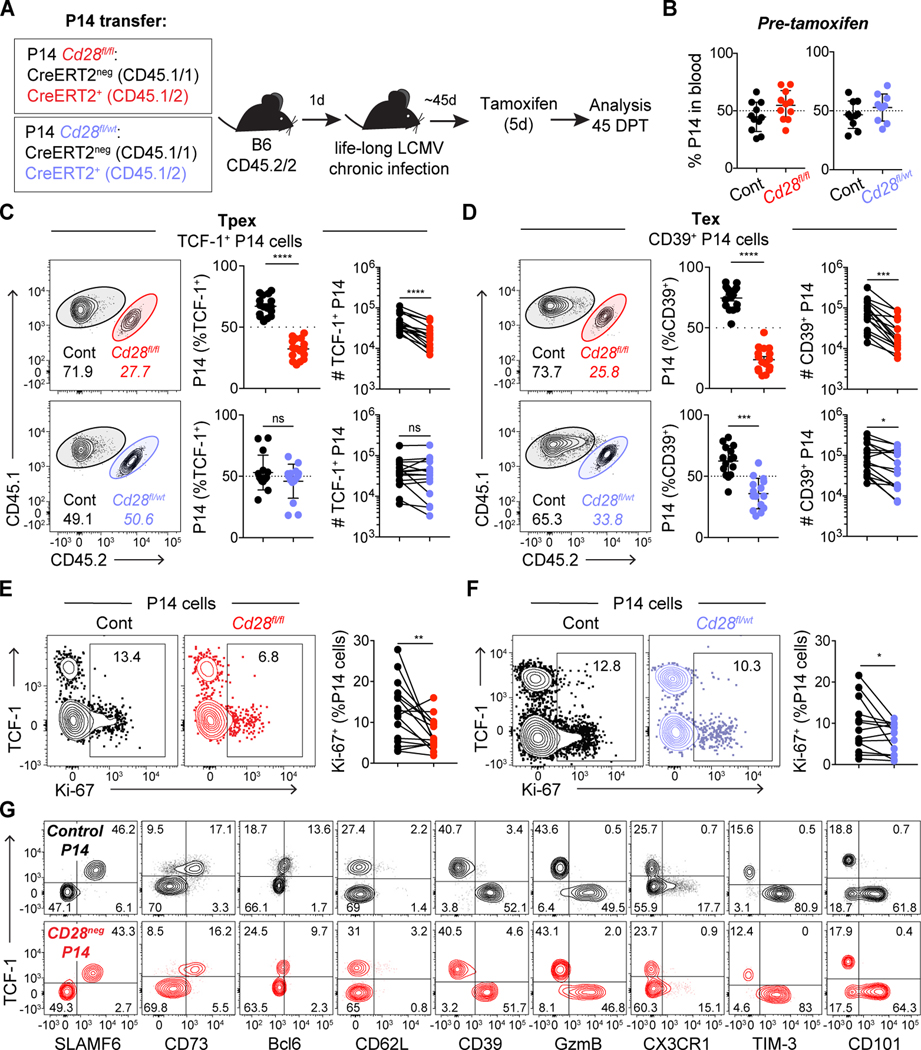
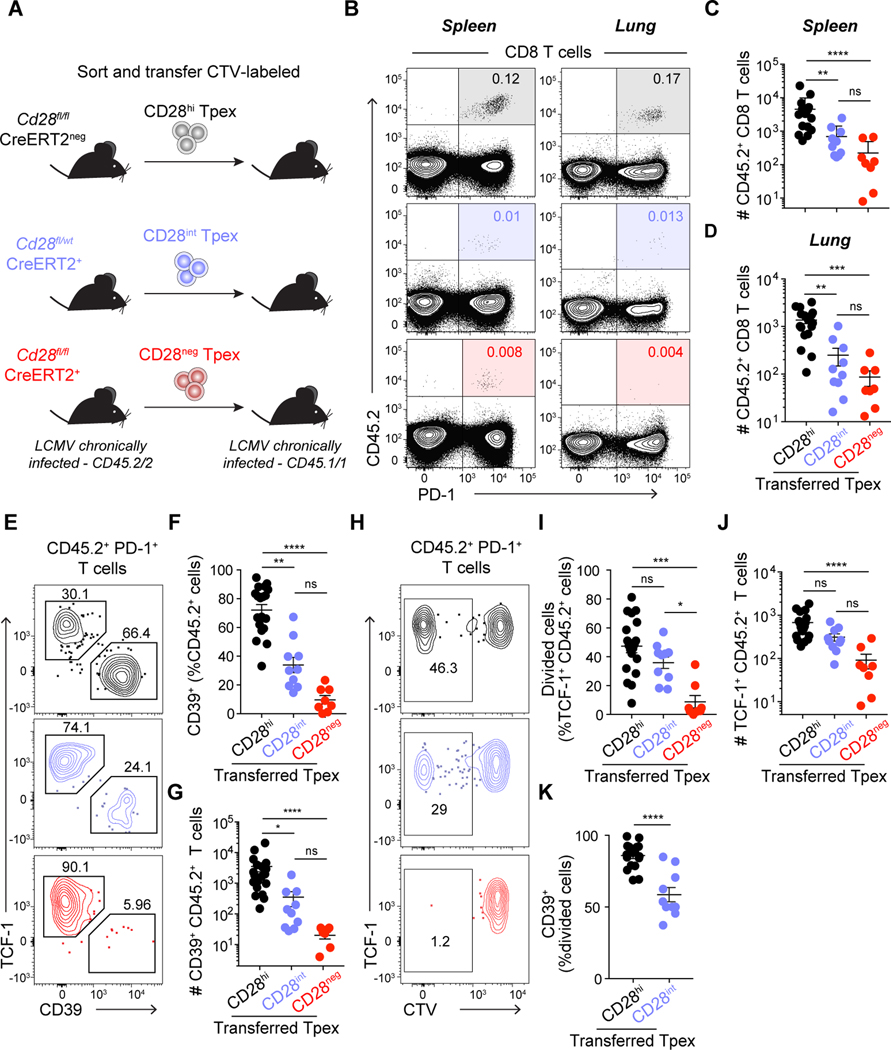
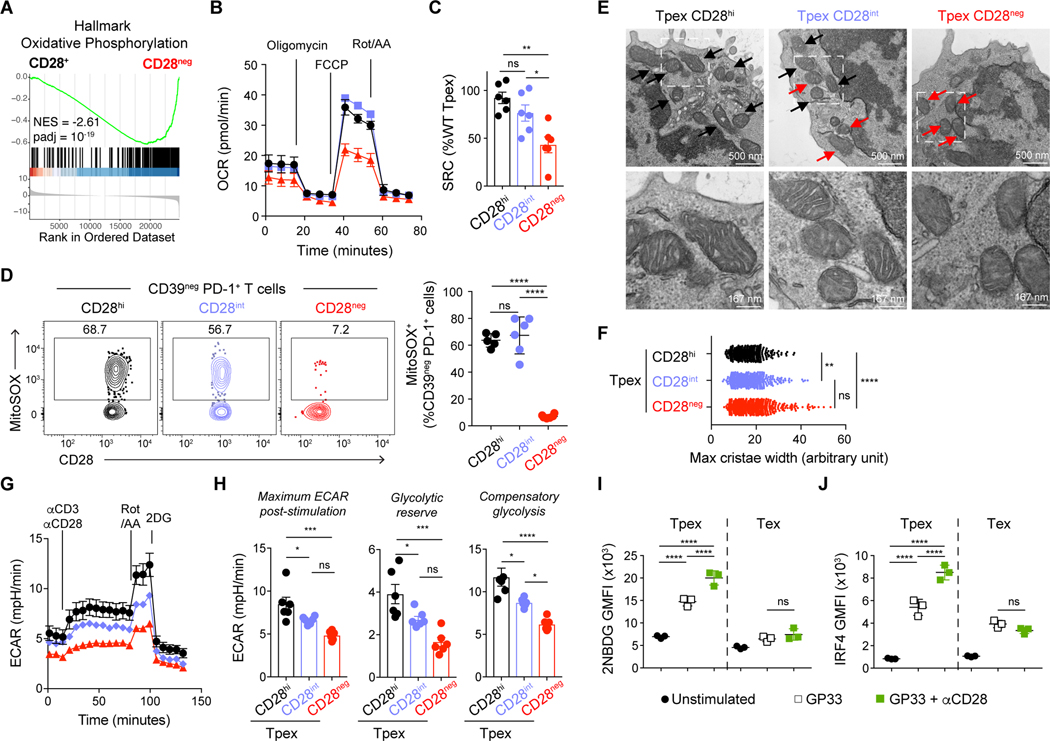
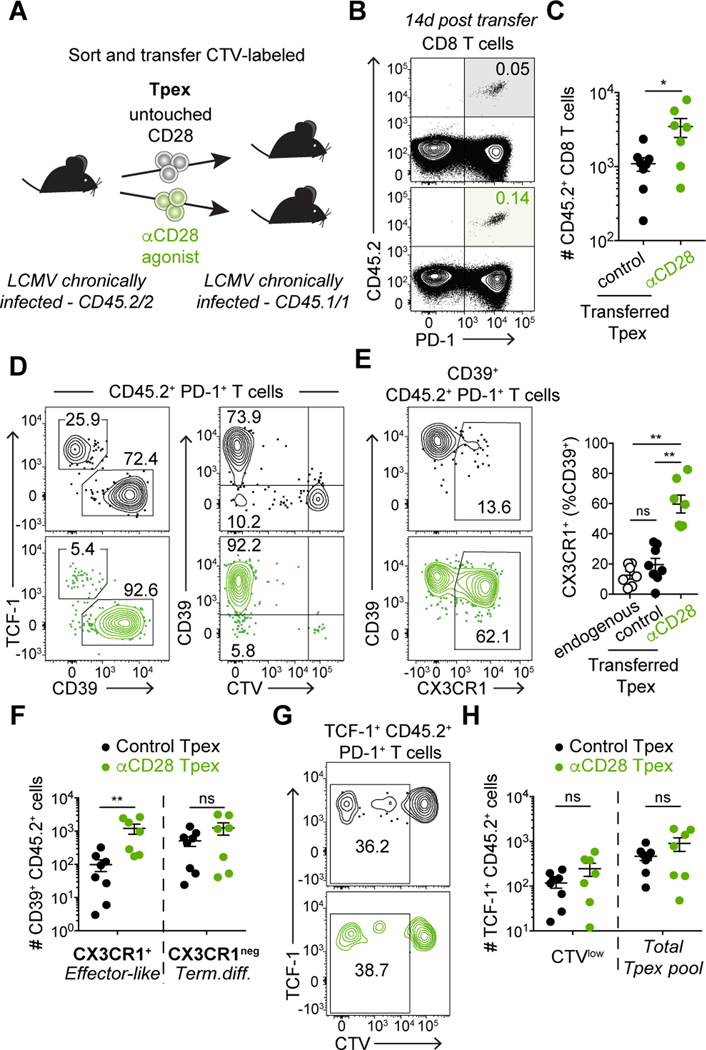
During persistent antigen stimulation, such as in chronic infections and cancer, CD8 T cells differentiate into a hypofunctional programmed death protein 1-positive (PD-1+) exhausted state. Exhausted CD8 T cell responses are maintained by precursors (Tpex) that express the transcription factor T cell factor 1 (TCF-1) and high levels of the costimulatory molecule CD28. Here, we demonstrate that sustained CD28 costimulation is required for maintenance of antiviral T cells during chronic infection. Low-level CD28 engagement preserved mitochondrial fitness and self-renewal of Tpex, whereas stronger CD28 signaling enhanced glycolysis and promoted Tpex differentiation into TCF-1neg exhausted CD8 T cells (Tex). Furthermore, enhanced differentiation by CD28 engagement did not reduce the Tpex pool. Together, these findings demonstrate that continuous CD28 engagement is needed to sustain PD-1+ CD8 T cells and suggest that increasing CD28 signaling promotes Tpex differentiation into more functional effector-like Tex, possibly without compromising long-term responses.
Conflict of interest statement
Competing interests:
The other authors declare that they have no competing interests.
Figures
References
-
- Hashimoto M, Kamphorst AO, Im SJ, Kissick HT, Pillai RN, Ramalingam SS, Araki K, Ahmed R, CD8 T Cell Exhaustion in Chronic Infection and Cancer: Opportunities for Interventions. Annu Rev Med 69, 301–318 (2018). - PubMed
-
- McLane LM, Abdel-Hakeem MS, Wherry EJ, CD8 T Cell Exhaustion During Chronic Viral Infection and Cancer. Annu Rev Immunol 37, 457–495 (2019). - PubMed
-
- Wu T, Ji Y, Moseman EA, Xu HC, Manglani M, Kirby M, Anderson SM, Handon R, Kenyon E, Elkahloun A, Wu W, Lang PA, Gattinoni L, McGavern DB, Schwartzberg PL, The TCF1-Bcl6 axis counteracts type I interferon to repress exhaustion and maintain T cell stemness. Sci Immunol 1, eaai8593 (2016). - PMC - PubMed
-
- Utzschneider DT, Charmoy M, Chennupati V, Pousse L, Ferreira DP, Calderon-Copete S, Danilo M, Alfei F, Hofmann M, Wieland D, Pradervand S, Thimme R, Zehn D, Held W, T Cell Factor 1-Expressing Memory-like CD8(+) T Cells Sustain the Immune Response to Chronic Viral Infections. Immunity 45, 415–427 (2016). - PubMed
-
- He R, Hou S, Liu C, Zhang A, Bai Q, Han M, Yang Y, Wei G, Shen T, Yang X, Xu L, Chen X, Hao Y, Wang P, Zhu C, Ou J, Liang H, Ni T, Zhang X, Zhou X, Deng K, Chen Y, Luo Y, Xu J, Qi H, Wu Y, Ye L, Follicular CXCR5- expressing CD8(+) T cells curtail chronic viral infection. Nature 537, 412–428 (2016). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
