The Importance of the "Time Factor" for the Evaluation of Inhibition Mechanisms: The Case of Selected HDAC6 Inhibitors
- PMID: 37626935
- PMCID: PMC10452033
- DOI: 10.3390/biology12081049
The Importance of the "Time Factor" for the Evaluation of Inhibition Mechanisms: The Case of Selected HDAC6 Inhibitors
Abstract
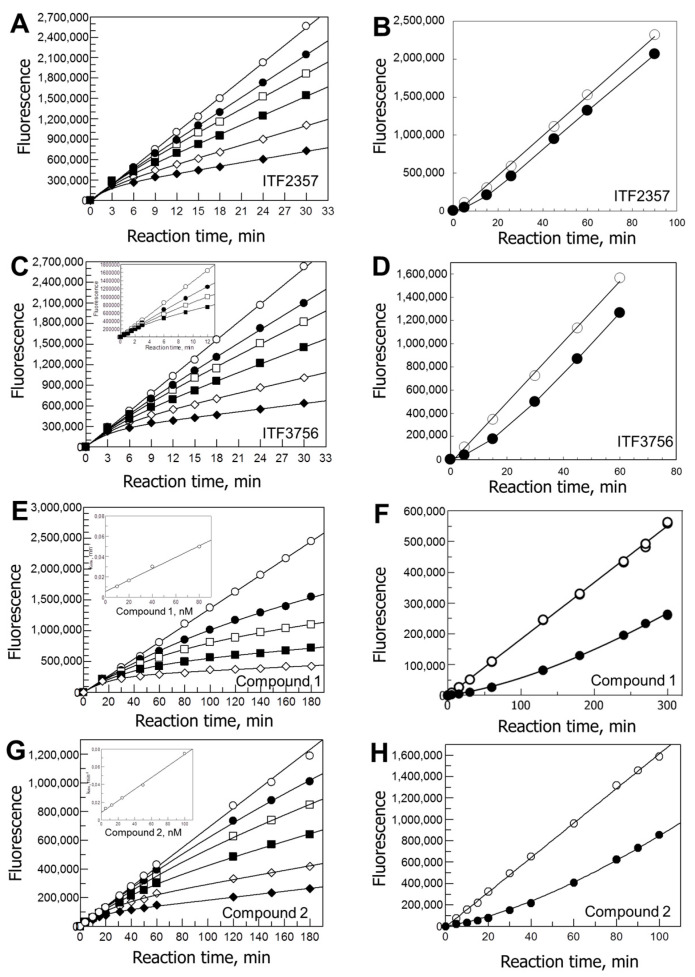
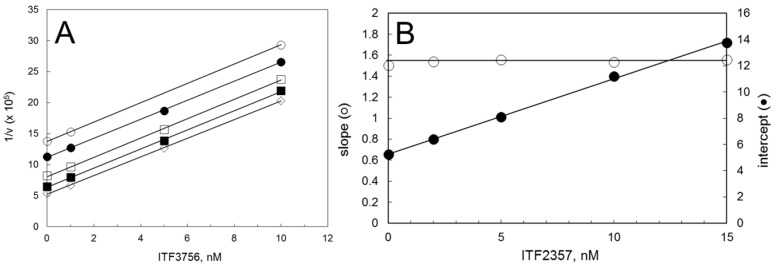
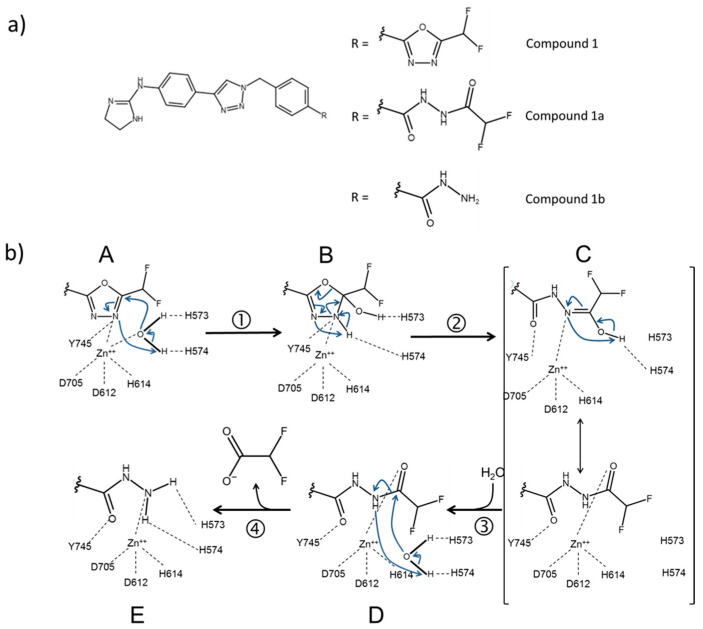
Histone deacetylases (HDACs) participate with histone acetyltransferases in the modulation of the biological activity of a broad array of proteins, besides histones. Histone deacetylase 6 is unique among HDAC as it contains two catalytic domains, an N-terminal microtubule binding region and a C-terminal ubiquitin binding domain. Most of its known biological roles are related to its protein lysine deacetylase activity in the cytoplasm. The design of specific inhibitors is the focus of a large number of medicinal chemistry programs in the academy and industry because lowering HDAC6 activity has been demonstrated to be beneficial for the treatment of several diseases, including cancer, and neurological and immunological disorders. Here, we show how re-evaluation of the mechanism of action of selected HDAC6 inhibitors, by monitoring the time-dependence of the onset and relief of the inhibition, revealed instances of slow-binding/slow-release inhibition. The same approach, in conjunction with X-ray crystallography, in silico modeling and mass spectrometry, helped to propose a model of inhibition of HDAC6 by a novel difluoromethyloxadiazole-based compound that was found to be a slow-binding substrate analog of HDAC6, giving rise to a tightly bound, long-lived inhibitory derivative.
Keywords: cancer; difluoromethyloxadiazoles; histone deacetylase; histone deacetylase 6; hydroxamates; immunomodulation; inhibition; kinetics; medicinal chemistry; neurodegeneration.
Conflict of interest statement
The authors have declared that no competing interest exists. Edoardo Cellupica, Gianluca Caprini, Gianluca Fossati, Paola Cordella, Mattia Marchini, Ilaria Rocchio, Giovanni Sandrone, Andrea Stevenazzi, Barbara Vergani, and Christian Steinkühler are employees of Italfarmaco SpA. The work in the laboratory of Maria Antonietta Vanoni was carried out with funds from Italfarmaco SpA that supported the training fellowship of Doris Mirdita.
Figures