

Modeling specific aneuploidies: from karyotype manipulations to biological insights
- PMID: 37640903
- PMCID: PMC10462580
- DOI: 10.1007/s10577-023-09735-7
Modeling specific aneuploidies: from karyotype manipulations to biological insights
Abstract
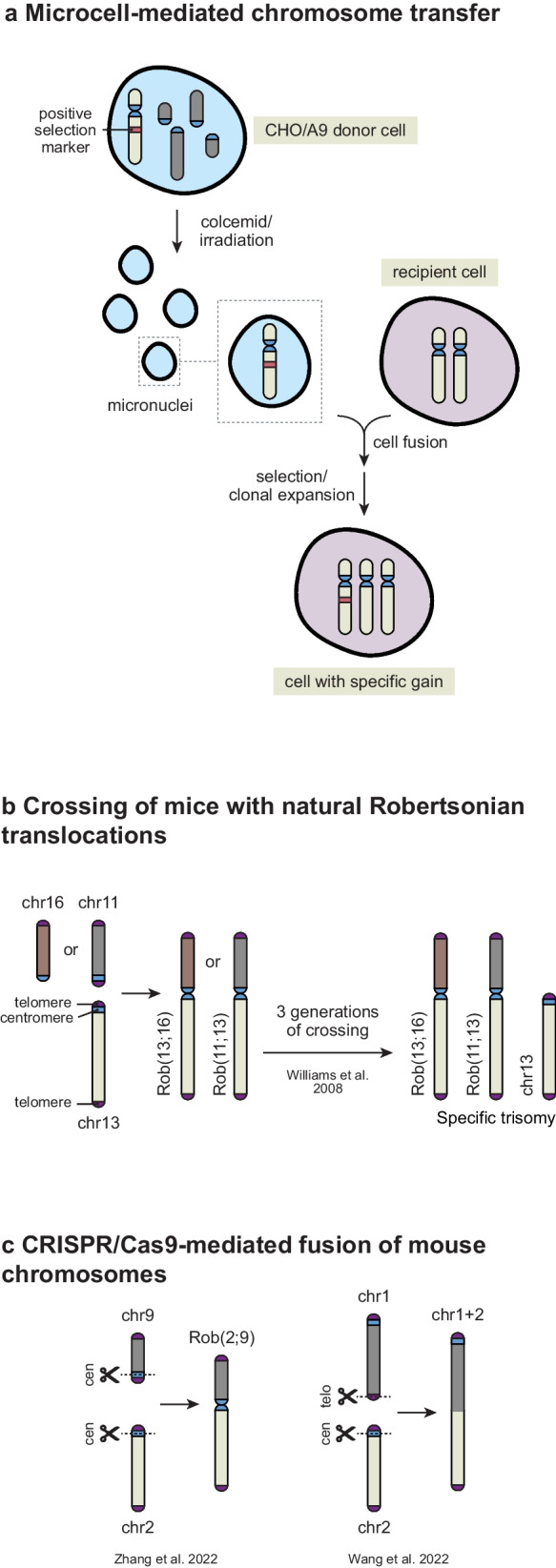
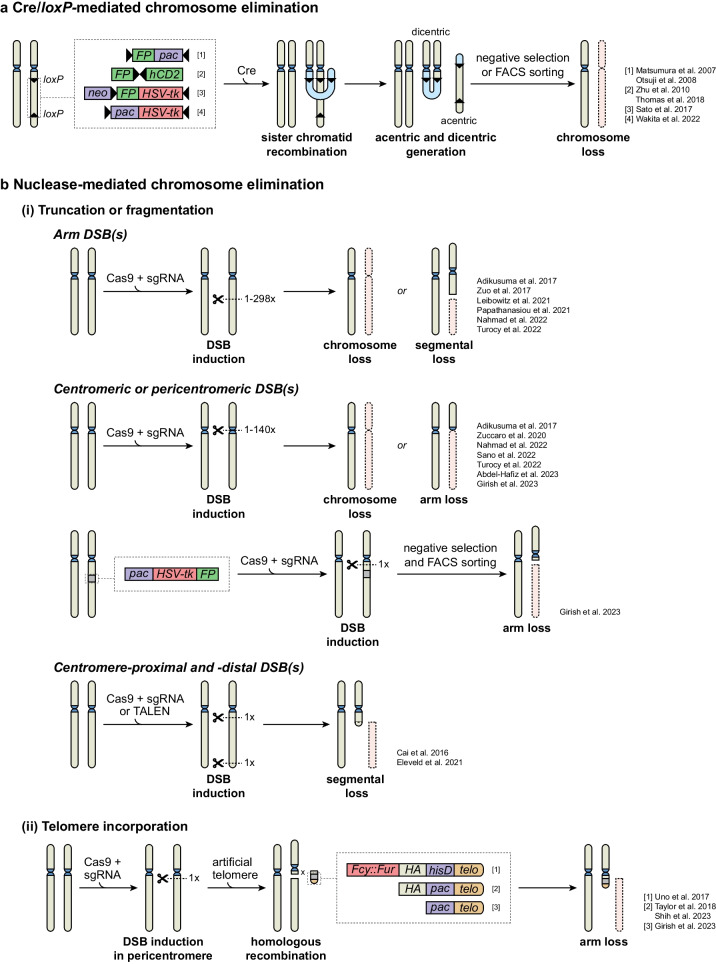
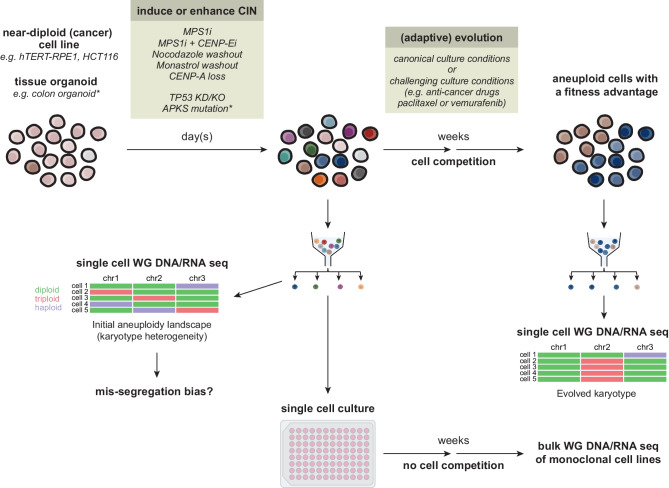
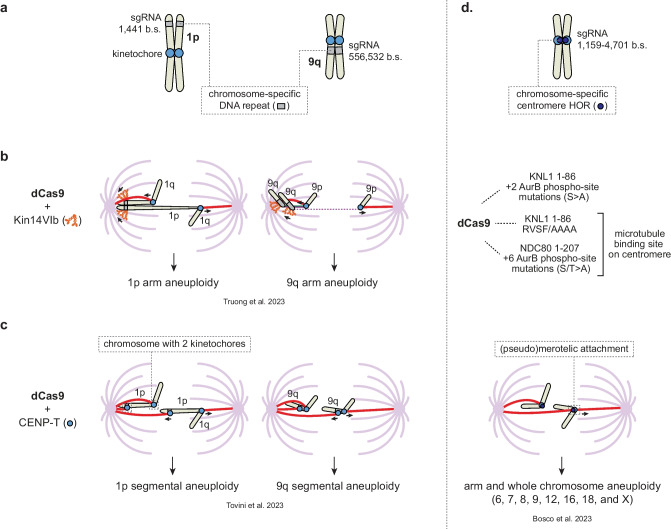
An abnormal chromosome number, or aneuploidy, underlies developmental disorders and is a common feature of cancer, with different cancer types exhibiting distinct patterns of chromosomal gains and losses. To understand how specific aneuploidies emerge in certain tissues and how they contribute to disease development, various methods have been developed to alter the karyotype of mammalian cells and mice. In this review, we provide an overview of both classic and novel strategies for inducing or selecting specific chromosomal gains and losses in human and murine cell systems. We highlight how these customized aneuploidy models helped expanding our knowledge of the consequences of specific aneuploidies to (cancer) cell physiology.
Keywords: Aneuploidy; CIN; CRISPR/Cas9; cancer; chromosome.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Clonal selection of stable aneuploidies in progenitor cells drives high-prevalence tumorigenesis.Genes Dev. 2021 Aug 1;35(15-16):1079-1092. doi: 10.1101/gad.348341.121. Epub 2021 Jul 15. Genes Dev. 2021. PMID: 34266888 Free PMC article.
-
Quantifying chromosomal instability from intratumoral karyotype diversity using agent-based modeling and Bayesian inference.Elife. 2022 Apr 5;11:e69799. doi: 10.7554/eLife.69799. Elife. 2022. PMID: 35380536 Free PMC article.
-
[Diagnosis of aneuploidy with fluorescence in situ hybridization (FISH); value in pregnancies with increased risk for chromosome aberrations].Z Geburtshilfe Neonatol. 2000 Jan-Feb;204(1):1-7. doi: 10.1055/s-2000-10188. Z Geburtshilfe Neonatol. 2000. PMID: 10721179 German.
-
Targeting chromosomal instability and aneuploidy in cancer.Trends Pharmacol Sci. 2024 Mar;45(3):210-224. doi: 10.1016/j.tips.2024.01.009. Epub 2024 Feb 13. Trends Pharmacol Sci. 2024. PMID: 38355324 Review.
-
Chromosome instability and aneuploidy in the mammalian brain.Chromosome Res. 2023 Nov 1;31(4):32. doi: 10.1007/s10577-023-09740-w. Chromosome Res. 2023. PMID: 37910282 Free PMC article. Review.
Cited by
-
Trisomic rescue via allele-specific multiple chromosome cleavage using CRISPR-Cas9 in trisomy 21 cells.PNAS Nexus. 2025 Feb 18;4(2):pgaf022. doi: 10.1093/pnasnexus/pgaf022. eCollection 2025 Feb. PNAS Nexus. 2025. PMID: 39967679 Free PMC article.
-
Patterns of Aneuploidy and Signaling Consequences in Cancer.Cancer Res. 2024 Aug 15;84(16):2575-2587. doi: 10.1158/0008-5472.CAN-24-0169. Cancer Res. 2024. PMID: 38924459 Free PMC article. Review.
-
Chromosome Transplantation: Opportunities and Limitations.Cells. 2024 Apr 11;13(8):666. doi: 10.3390/cells13080666. Cells. 2024. PMID: 38667281 Free PMC article. Review.
-
Aneuploidy as a driver of human cancer.Nat Genet. 2024 Oct;56(10):2014-2026. doi: 10.1038/s41588-024-01916-2. Epub 2024 Oct 2. Nat Genet. 2024. PMID: 39358600 Review.
-
Deletion of 17p in cancers: Guilt by (p53) association.Oncogene. 2025 Mar;44(10):637-651. doi: 10.1038/s41388-025-03300-8. Epub 2025 Feb 18. Oncogene. 2025. PMID: 39966556 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
