

Propagation of activity through the cortical hierarchy and perception are determined by neural variability
- PMID: 37640911
- PMCID: PMC10471496
- DOI: 10.1038/s41593-023-01413-5
Propagation of activity through the cortical hierarchy and perception are determined by neural variability
Abstract
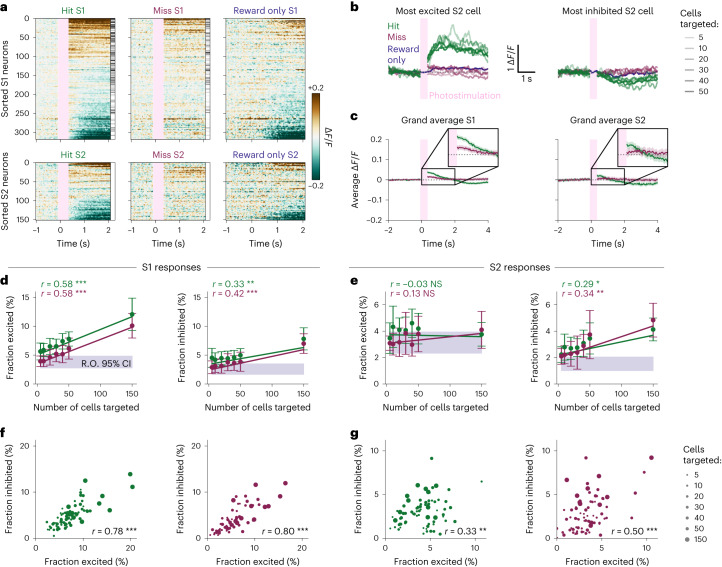
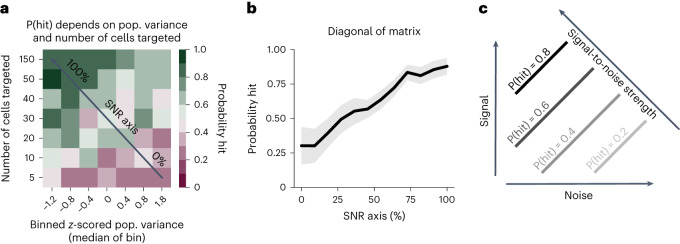
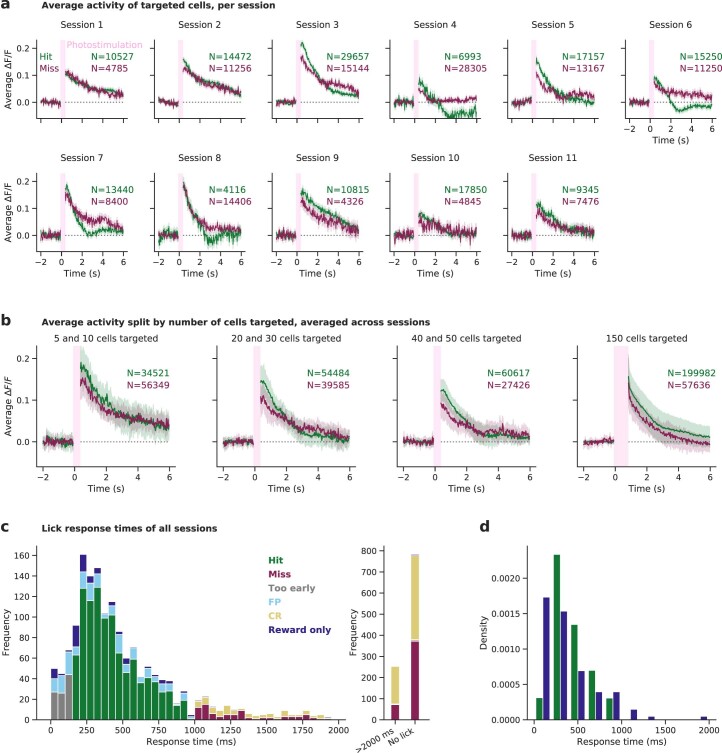
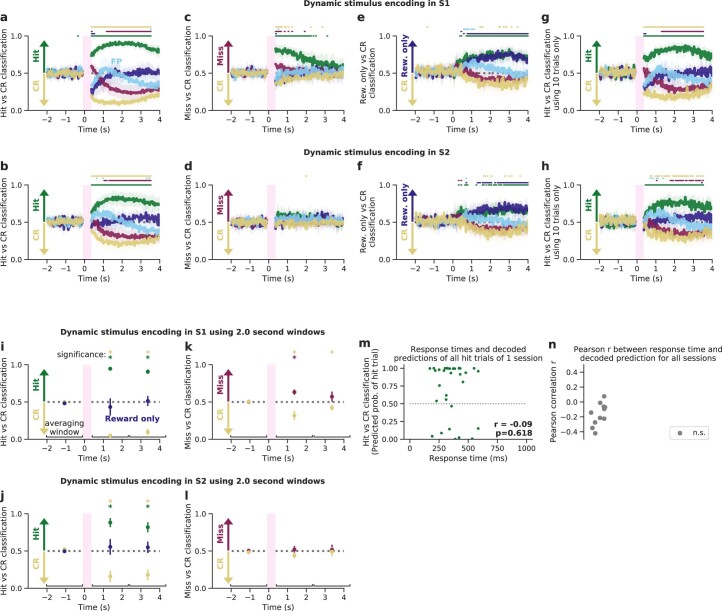
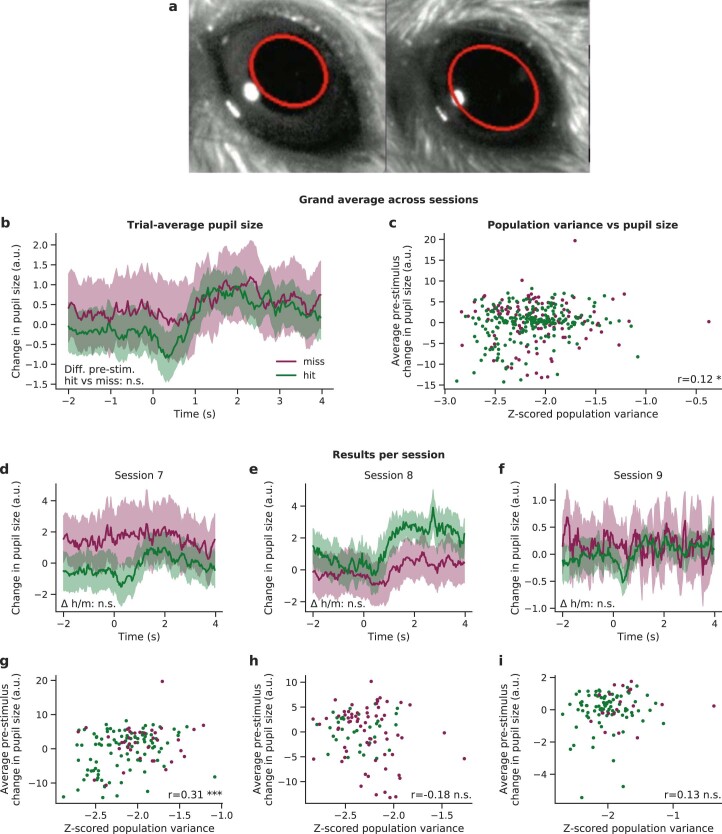
Brains are composed of anatomically and functionally distinct regions performing specialized tasks, but regions do not operate in isolation. Orchestration of complex behaviors requires communication between brain regions, but how neural dynamics are organized to facilitate reliable transmission is not well understood. Here we studied this process directly by generating neural activity that propagates between brain regions and drives behavior, assessing how neural populations in sensory cortex cooperate to transmit information. We achieved this by imaging two densely interconnected regions-the primary and secondary somatosensory cortex (S1 and S2)-in mice while performing two-photon photostimulation of S1 neurons and assigning behavioral salience to the photostimulation. We found that the probability of perception is determined not only by the strength of the photostimulation but also by the variability of S1 neural activity. Therefore, maximizing the signal-to-noise ratio of the stimulus representation in cortex relative to the noise or variability is critical to facilitate activity propagation and perception.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures











Similar articles
-
Multielectrode Recordings in the Somatosensory System.In: Nicolelis MAL, editor. Methods for Neural Ensemble Recordings. 2nd edition. Boca Raton (FL): CRC Press/Taylor & Francis; 2008. Chapter 6. In: Nicolelis MAL, editor. Methods for Neural Ensemble Recordings. 2nd edition. Boca Raton (FL): CRC Press/Taylor & Francis; 2008. Chapter 6. PMID: 21204443 Free Books & Documents. Review.
-
Context-Dependent Sensory Processing across Primary and Secondary Somatosensory Cortex.Neuron. 2020 May 6;106(3):515-525.e5. doi: 10.1016/j.neuron.2020.02.004. Epub 2020 Mar 11. Neuron. 2020. PMID: 32164873 Free PMC article.
-
Ipsilateral Stimulus Encoding in Primary and Secondary Somatosensory Cortex of Awake Mice.J Neurosci. 2022 Mar 30;42(13):2701-2715. doi: 10.1523/JNEUROSCI.1417-21.2022. Epub 2022 Feb 8. J Neurosci. 2022. PMID: 35135855 Free PMC article.
-
Neocortical dynamics during whisker-based sensory discrimination in head-restrained mice.Neuroscience. 2018 Jan 1;368:57-69. doi: 10.1016/j.neuroscience.2017.09.003. Epub 2017 Sep 14. Neuroscience. 2018. PMID: 28919043 Free PMC article. Review.
-
Behaviorally relevant decision coding in primary somatosensory cortex neurons.Nat Neurosci. 2022 Sep;25(9):1225-1236. doi: 10.1038/s41593-022-01151-0. Epub 2022 Aug 30. Nat Neurosci. 2022. PMID: 36042310 Free PMC article.
Cited by
-
Touch-evoked traveling waves establish a translaminar spacetime code.Sci Adv. 2025 Jan 31;11(5):eadr4038. doi: 10.1126/sciadv.adr4038. Epub 2025 Jan 31. Sci Adv. 2025. PMID: 39889002 Free PMC article.
-
Alpha-band fluctuations represent behaviorally relevant excitability changes as a consequence of top-down guided spatial attention in a probabilistic spatial cueing design.Imaging Neurosci (Camb). 2024 Oct 15;2:imag-2-00312. doi: 10.1162/imag_a_00312. eCollection 2024. Imaging Neurosci (Camb). 2024. PMID: 40800404 Free PMC article.
-
What does the mean mean? A simple test for neuroscience.PLoS Comput Biol. 2024 Apr 19;20(4):e1012000. doi: 10.1371/journal.pcbi.1012000. eCollection 2024 Apr. PLoS Comput Biol. 2024. PMID: 38640119 Free PMC article.
-
Neurodegenerative disease of the brain: a survey of interdisciplinary approaches.J R Soc Interface. 2023 Jan;20(198):20220406. doi: 10.1098/rsif.2022.0406. Epub 2023 Jan 18. J R Soc Interface. 2023. PMID: 36651180 Free PMC article. Review.
-
In Vivo Two-Photon Microscopy Reveals Sensory-Evoked Serotonin (5-HT) Release in Adult Mammalian Neocortex.ACS Chem Neurosci. 2024 Feb 7;15(3):456-461. doi: 10.1021/acschemneuro.3c00725. Epub 2024 Jan 22. ACS Chem Neurosci. 2024. PMID: 38251903 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
