

Schwann cells are axo-protective after injury irrespective of myelination status in mouse Schwann cell-neuron cocultures
- PMID: 37642648
- PMCID: PMC10546878
- DOI: 10.1242/jcs.261557
Schwann cells are axo-protective after injury irrespective of myelination status in mouse Schwann cell-neuron cocultures
Abstract
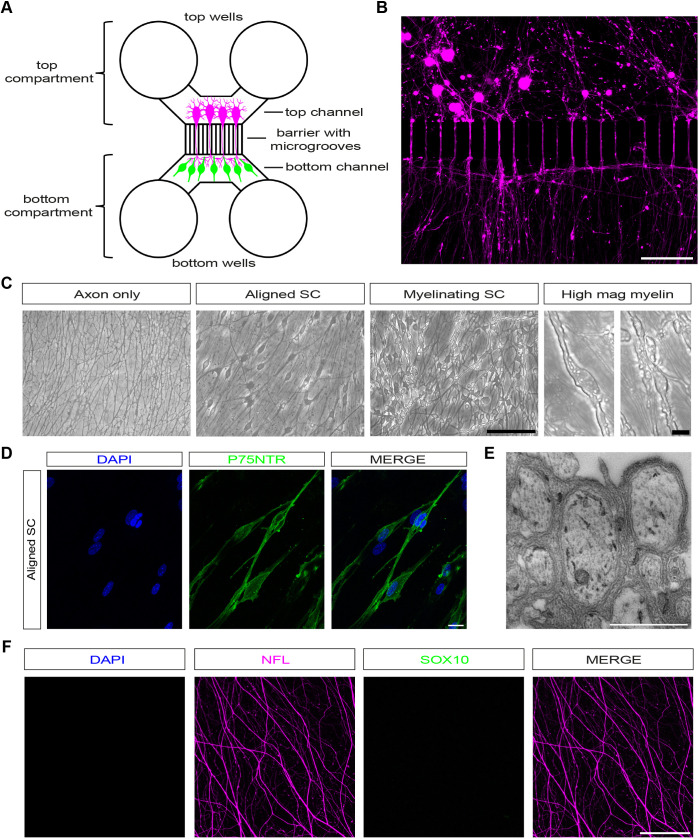
Myelinating Schwann cell (SC)-dorsal root ganglion (DRG) neuron cocultures are an important technique for understanding cell-cell signalling and interactions during peripheral nervous system (PNS) myelination, injury, and regeneration. Although methods using rat SCs and neurons or mouse DRG explants are commonplace, there are no established protocols for compartmentalised myelinating cocultures with dissociated mouse cells. There consequently is a need for a coculture protocol that allows separate genetic manipulation of mouse SCs or neurons, or use of cells from different transgenic animals to complement in vivo mouse experiments. However, inducing myelination of dissociated mouse SCs in culture is challenging. Here, we describe a new method to coculture dissociated mouse SCs and DRG neurons in microfluidic chambers and induce robust myelination. Cocultures can be axotomised to study injury and used for drug treatments, and cells can be lentivirally transduced for live imaging. We used this model to investigate axon degeneration after traumatic axotomy and find that SCs, irrespective of myelination status, are axo-protective. At later timepoints after injury, live imaging of cocultures shows that SCs break up, ingest and clear axonal debris.
Keywords: Axon degeneration; Coculture; Dorsal root ganglion neuron; Mouse; Myelination; Schwann cell; Wallerian degeneration.
© 2023. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests M.P.C. is a consultant for NuraBio. The remaining authors declare no competing interests.
Figures



Similar articles
-
In Vitro Myelination of Peripheral Axons in a Coculture of Rat Dorsal Root Ganglion Explants and Schwann Cells.J Vis Exp. 2023 Feb 10;(192). doi: 10.3791/64768. J Vis Exp. 2023. PMID: 36847366
-
Establishing Myelinating Cocultures Using Human iPSC-Derived Sensory Neurons to Investigate Axonal Degeneration and Demyelination.Methods Mol Biol. 2020;2143:111-129. doi: 10.1007/978-1-0716-0585-1_9. Methods Mol Biol. 2020. PMID: 32524476
-
DRG Neuron/Schwann Cells Myelinating Cocultures.Methods Mol Biol. 2018;1791:115-129. doi: 10.1007/978-1-4939-7862-5_9. Methods Mol Biol. 2018. PMID: 30006705
-
Cadm3 (Necl-1) interferes with the activation of the PI3 kinase/Akt signaling cascade and inhibits Schwann cell myelination in vitro.Glia. 2016 Dec;64(12):2247-2262. doi: 10.1002/glia.23072. Epub 2016 Sep 23. Glia. 2016. PMID: 27658374 Free PMC article.
-
Nerve impulses regulate myelination through purinergic signalling.Novartis Found Symp. 2006;276:148-58; discussion 158-61, 233-7, 275-81. Novartis Found Symp. 2006. PMID: 16805428 Review.
Cited by
-
Influence of Magnesium Degradation on Schwannoma Cell Responses to Nerve Injury Using an In Vitro Injury Model.J Funct Biomater. 2024 Mar 31;15(4):88. doi: 10.3390/jfb15040088. J Funct Biomater. 2024. PMID: 38667545 Free PMC article.
References
-
- Arthur-Farraj, P. J., Latouche, M., Wilton, D. K., Quintes, S., Chabrol, E., Banerjee, A., Woodhoo, A., Jenkins, B., Rahman, M., Turmaine, M.et al. (2012). c-Jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron 75, 633-647. 10.1016/j.neuron.2012.06.021 - DOI - PMC - PubMed
-
- Arthur-Farraj, P. J., Morgan, C. C., Adamowicz, M., Gomez-Sanchez, J. A., Fazal, S. V., Beucher, A., Razzaghi, B., Mirsky, R., Jessen, K. R. and Aitman, T. J. (2017). Changes in the coding and non-coding transcriptome and DNA methylome that define the Schwann cell repair phenotype after nerve injury. Cell Rep. 20, 2719-2734. 10.1016/j.celrep.2017.08.064 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
