

This is a preprint.
Structural elucidation of recombinant Trichomonas vaginalis 20S proteasome bound to covalent inhibitors
- PMID: 37645851
- PMCID: PMC10462138
- DOI: 10.1101/2023.08.17.553660
Structural elucidation of recombinant Trichomonas vaginalis 20S proteasome bound to covalent inhibitors
Update in
-
Structural elucidation of recombinant Trichomonas vaginalis 20S proteasome bound to covalent inhibitors.Nat Commun. 2024 Oct 4;15(1):8621. doi: 10.1038/s41467-024-53022-w. Nat Commun. 2024. PMID: 39366995 Free PMC article.
Abstract
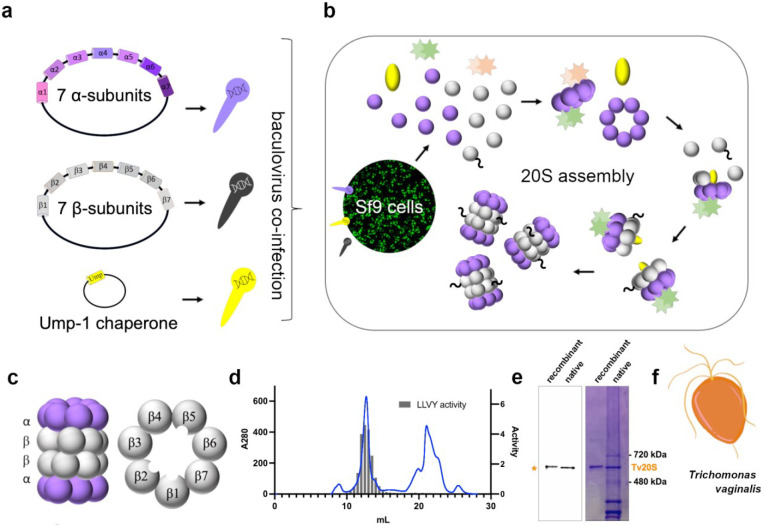
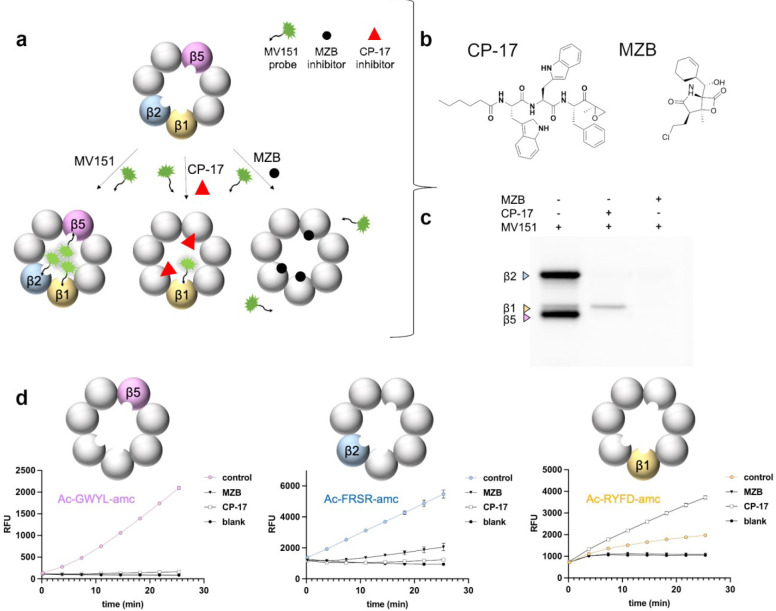
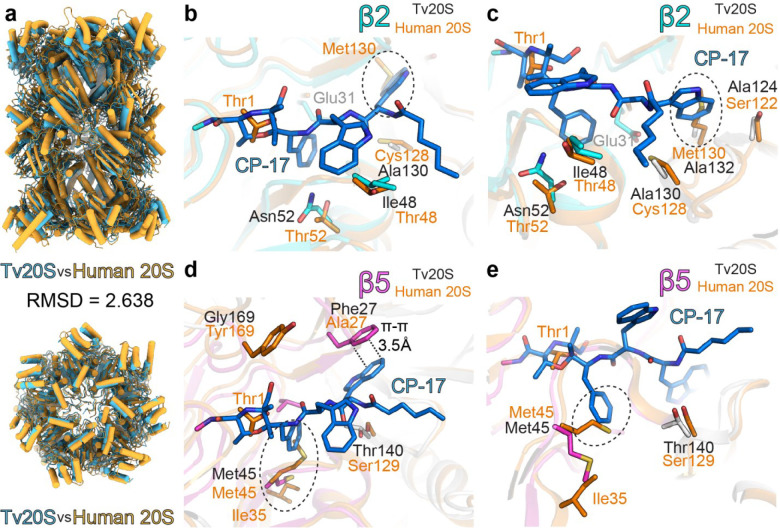
Proteasomes are essential for protein homeostasis in mammalian cells1-4 and in protozoan parasites such as Trichomonas vaginalis (Tv).5 Tv and other protozoan 20S proteasomes have been validated as druggable targets.6-8 However, in the case of Tv 20S proteasome (Tv20S), biochemical and structural studies were impeded by low yields and purity of the native proteasome. We successfully made recombinant Tv20S by expressing all seven α and seven β subunits together with the Ump-1 chaperone in insect cells. We isolated recombinant proteasome and showed that it was biochemically indistinguishable from the native enzyme. We confirmed that the recombinant Tv20S is inhibited by the natural product marizomib (MZB)9 and the recently developed peptide inhibitor carmaphycin-17 (CP-17)8,10. Specifically, MZB binds to the β1, β2 and β5 subunits, while CP-17 binds the β2 and β5 subunits. Next, we obtained cryo-EM structures of Tv20S in complex with these covalent inhibitors at 2.8Å resolution. The structures revealed the overall fold of the Tv20S and the binding mode of MZB and CP-17. Our work explains the low specificity of MZB and higher specificity of CP-17 towards Tv20S as compared to human proteasome and provides the platform for the development of Tv20S inhibitors for treatment of trichomoniasis.
Conflict of interest statement
Conflict of Interests The authors declare no conflict of interests.
Figures


References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous