

This is a preprint.
Population coding of time-varying sounds in the non-lemniscal Inferior Colliculus
- PMID: 37645904
- PMCID: PMC10461978
- DOI: 10.1101/2023.08.14.553263
Population coding of time-varying sounds in the non-lemniscal Inferior Colliculus
Update in
-
Population coding of time-varying sounds in the nonlemniscal inferior colliculus.J Neurophysiol. 2024 May 1;131(5):842-864. doi: 10.1152/jn.00013.2024. Epub 2024 Mar 20. J Neurophysiol. 2024. PMID: 38505907 Free PMC article.
Abstract
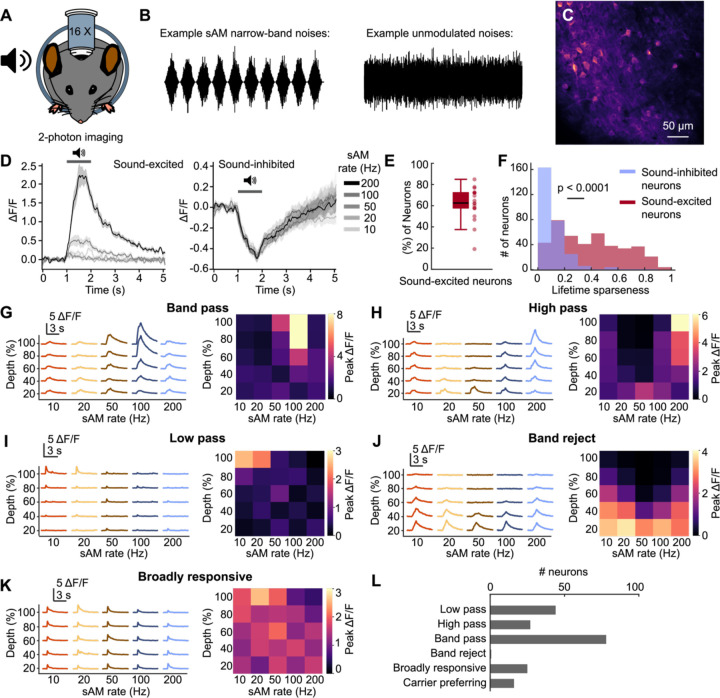
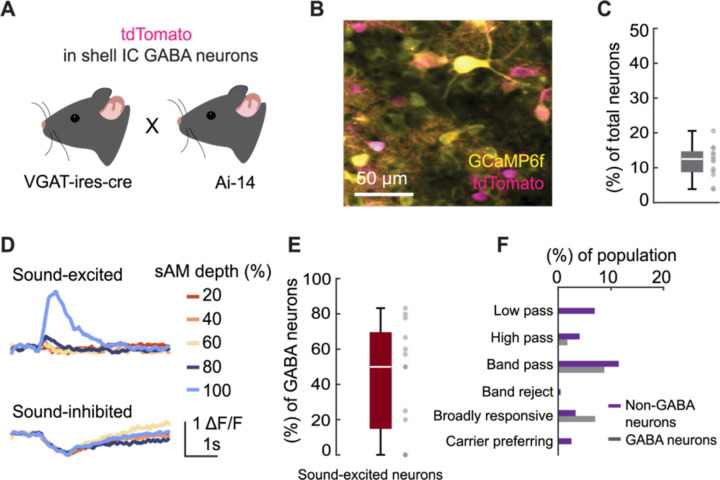
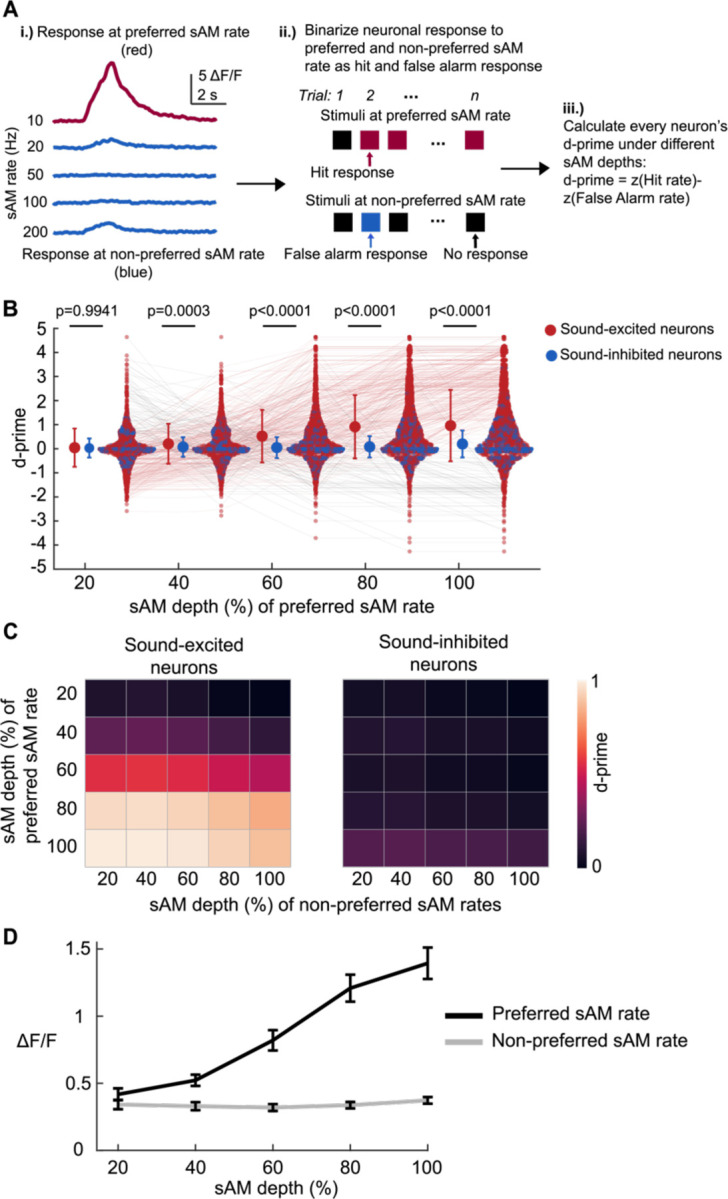
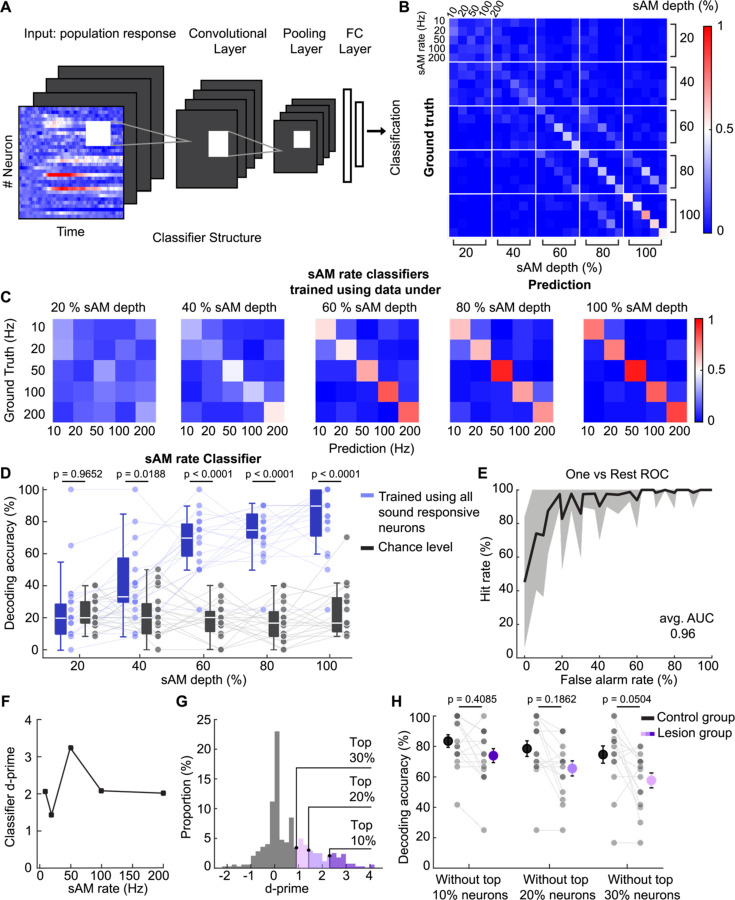
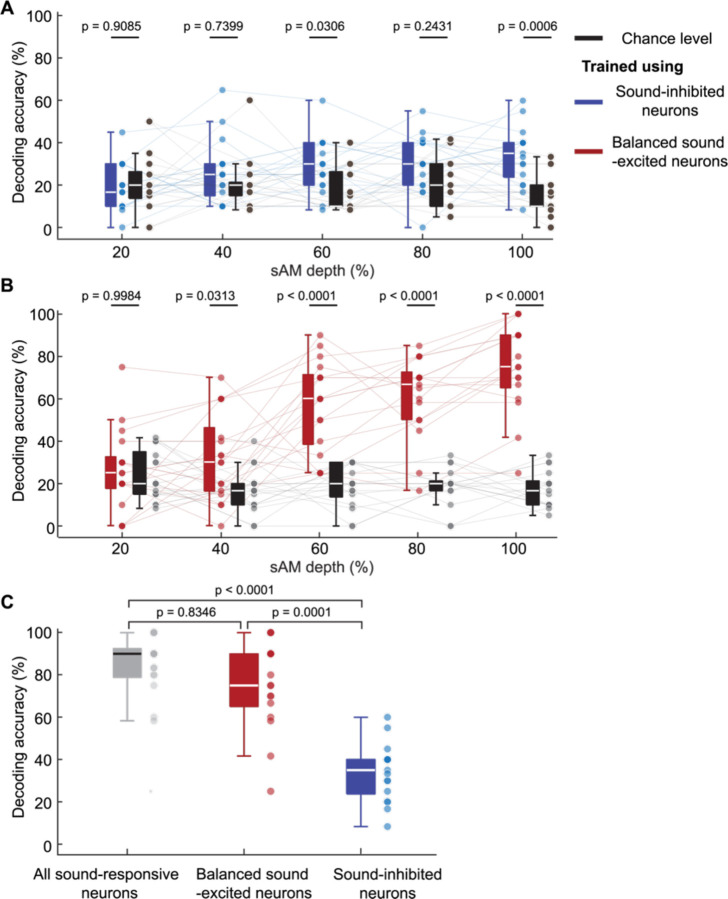
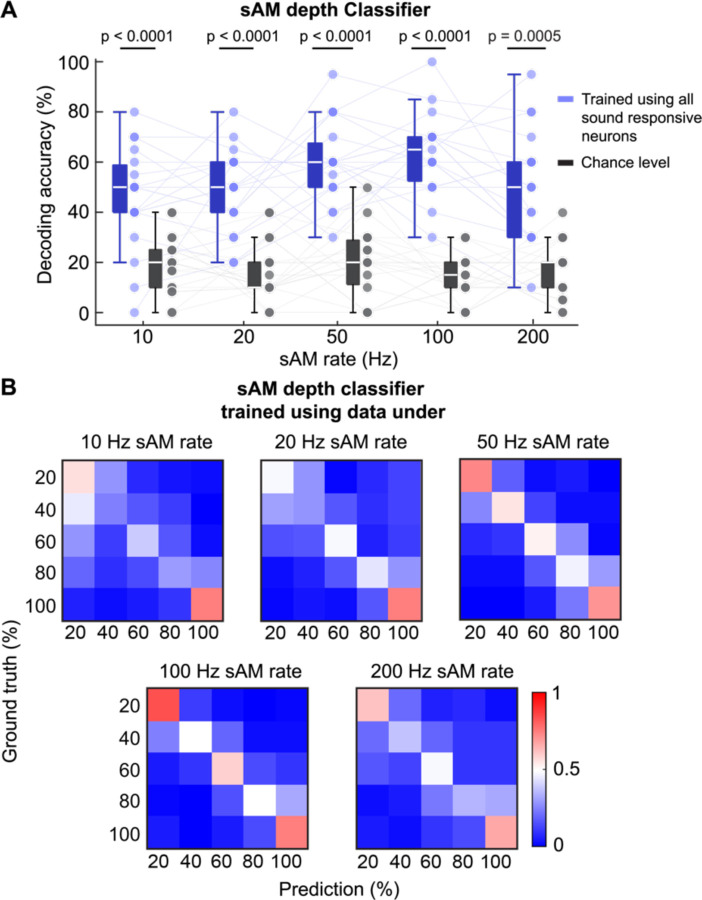
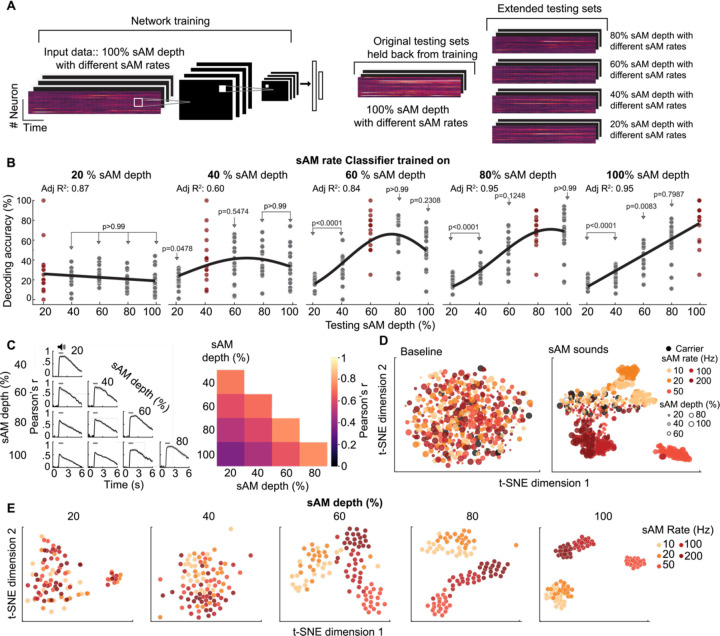
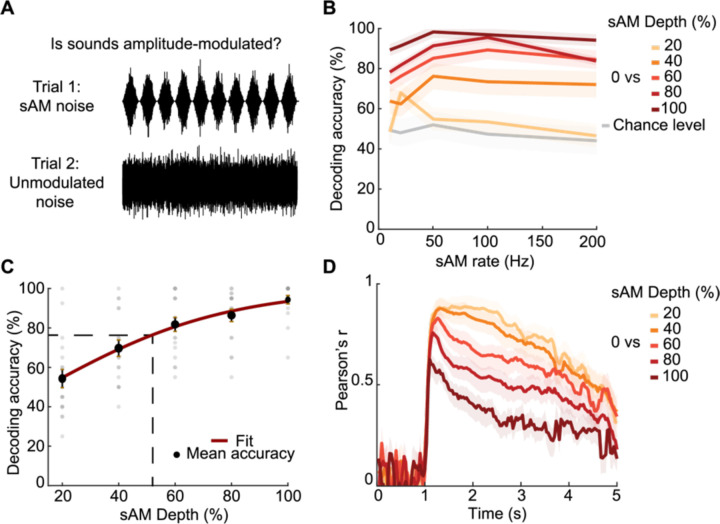
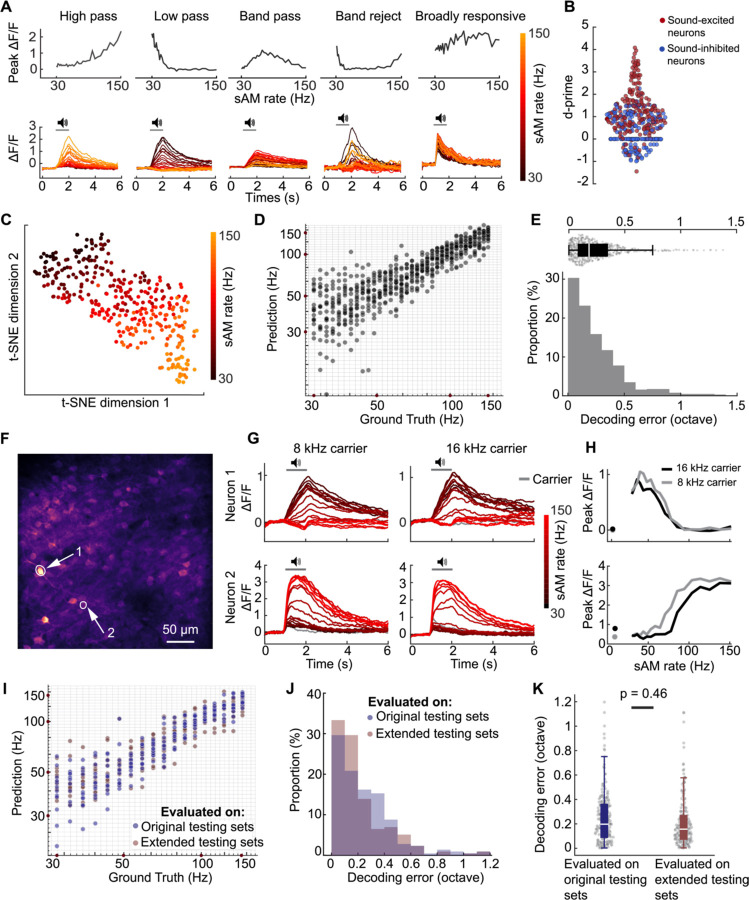
The inferior colliculus (IC) of the midbrain is important for complex sound processing, such as discriminating conspecific vocalizations and human speech. The IC's non-lemniscal, dorsal "shell" region is likely important for this process, as neurons in these layers project to higher-order thalamic nuclei that subsequently funnel acoustic signals to the amygdala and non-primary auditory cortices; forebrain circuits important for vocalization coding in a variety of mammals, including humans. However, the extent to which shell IC neurons transmit acoustic features necessary to discern vocalizations is less clear, owing to the technical difficulty of recording from neurons in the IC's superficial layers via traditional approaches. Here we use 2-photon Ca2+ imaging in mice of either sex to test how shell IC neuron populations encode the rate and depth of amplitude modulation, important sound cues for speech perception. Most shell IC neurons were broadly tuned, with a low neurometric discrimination of amplitude modulation rate; only a subset were highly selective to specific modulation rates. Nevertheless, neural network classifier trained on fluorescence data from shell IC neuron populations accurately classified amplitude modulation rate, and decoding accuracy was only marginally reduced when highly tuned neurons were omitted from training data. Rather, classifier accuracy increased monotonically with the modulation depth of the training data, such that classifiers trained on full-depth modulated sounds had median decoding errors of ~0.2 octaves. Thus, shell IC neurons may transmit time-varying signals via a population code, with perhaps limited reliance on the discriminative capacity of any individual neuron.
Conflict of interest statement
Conflict of interest statement: The authors report no competing interests.
Figures
Similar articles
-
Population coding of time-varying sounds in the nonlemniscal inferior colliculus.J Neurophysiol. 2024 May 1;131(5):842-864. doi: 10.1152/jn.00013.2024. Epub 2024 Mar 20. J Neurophysiol. 2024. PMID: 38505907 Free PMC article.
-
Mixed representations of sound and action in the auditory midbrain.bioRxiv [Preprint]. 2023 Sep 19:2023.09.19.558449. doi: 10.1101/2023.09.19.558449. bioRxiv. 2023. Update in: J Neurosci. 2024 Jul 24;44(30):e1831232024. doi: 10.1523/JNEUROSCI.1831-23.2024. PMID: 37786676 Free PMC article. Updated. Preprint.
-
Mixed Representations of Sound and Action in the Auditory Midbrain.J Neurosci. 2024 Jul 24;44(30):e1831232024. doi: 10.1523/JNEUROSCI.1831-23.2024. J Neurosci. 2024. PMID: 38918064 Free PMC article.
-
Circuits for processing dynamic interaural intensity disparities in the inferior colliculus.Hear Res. 2012 Jun;288(1-2):47-57. doi: 10.1016/j.heares.2012.01.011. Epub 2012 Feb 8. Hear Res. 2012. PMID: 22343068 Free PMC article. Review.
-
How do auditory cortex neurons represent communication sounds?Hear Res. 2013 Nov;305:102-12. doi: 10.1016/j.heares.2013.03.011. Epub 2013 Apr 17. Hear Res. 2013. PMID: 23603138 Review.
References
-
- Aitkin L, Tran L, Syka J (1994) The responses of neurons in subdivisions of the inferior colliculus of cats to tonal, noise and vocal stimuli. Exp Brain Res 98:53–64. - PubMed
-
- Aitkin LM, Webster WR, Veale JL, Crosby DC (1975) Inferior colliculus. I. Comparison of response properties of neurons in central, pericentral, and external nuclei of adult cat. J Neurophysiol 38:1196–1207. - PubMed
-
- Averbeck BB, Latham PE, Pouget A (2006) Neural correlations, population coding and computation. Nat Rev Neurosci 7:358–66. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous