

This is a preprint.
Mechanical force regulates ligand binding and function of PD-1
- PMID: 37645980
- PMCID: PMC10462004
- DOI: 10.1101/2023.08.13.553152
Mechanical force regulates ligand binding and function of PD-1
Update in
-
Mechanical force regulates ligand binding and function of PD-1.Nat Commun. 2024 Sep 27;15(1):8339. doi: 10.1038/s41467-024-52565-2. Nat Commun. 2024. PMID: 39333505 Free PMC article.
Abstract
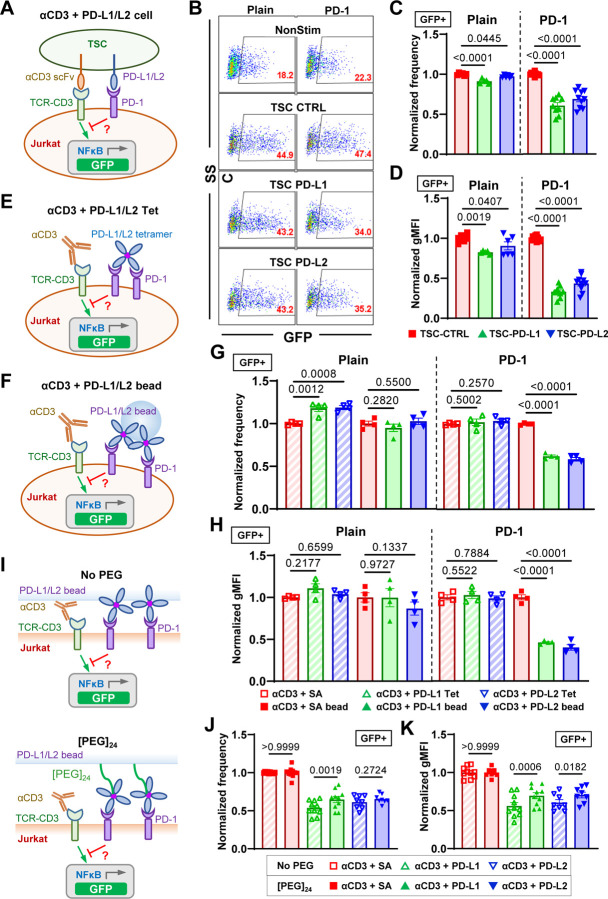
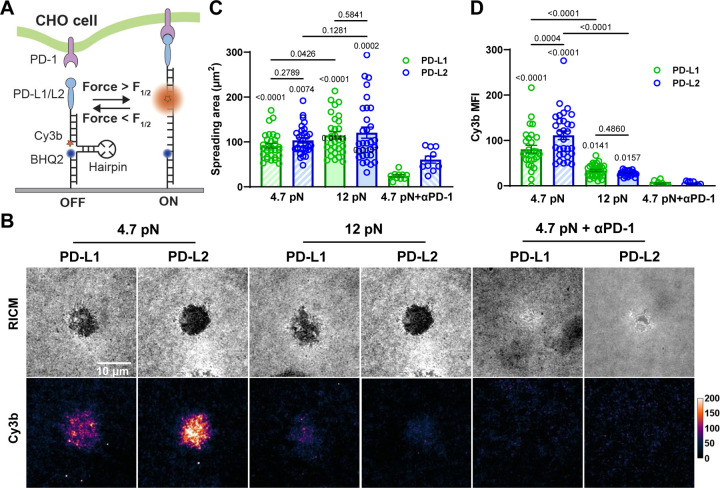
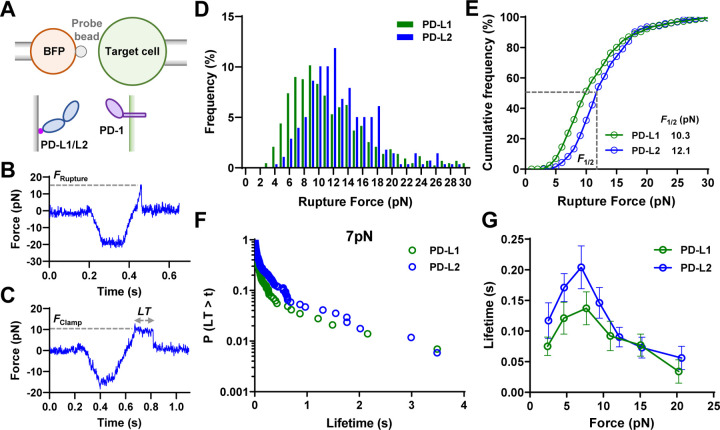
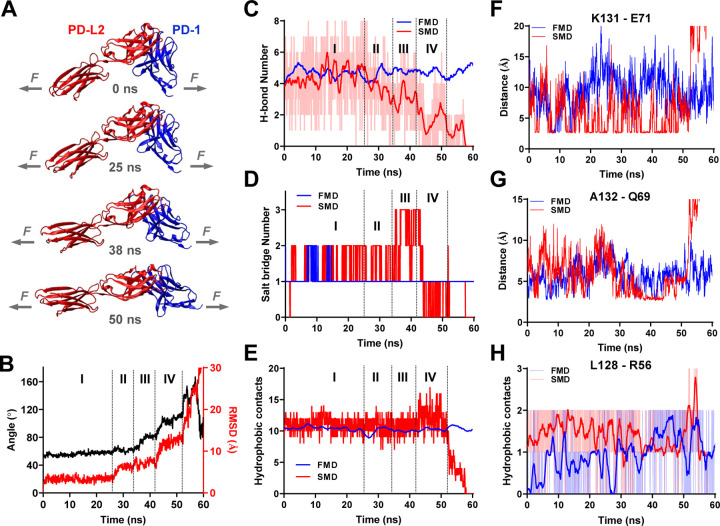
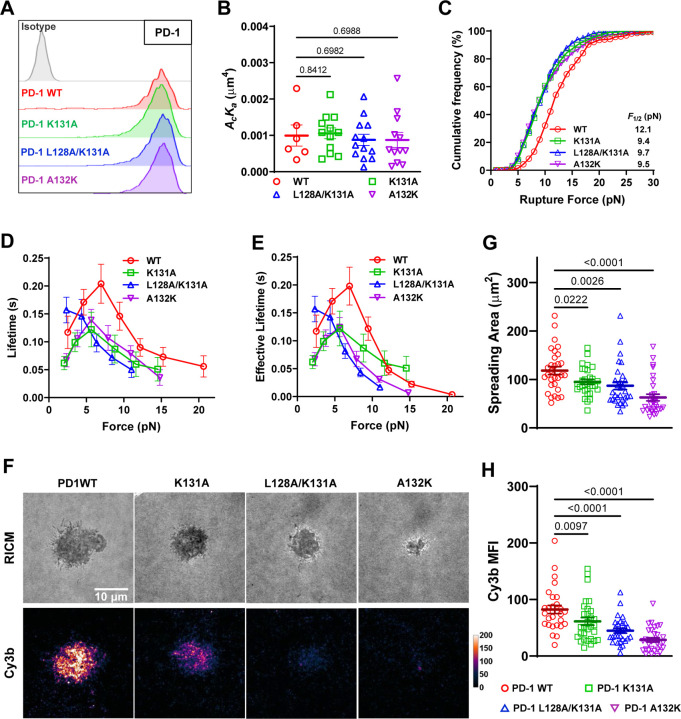
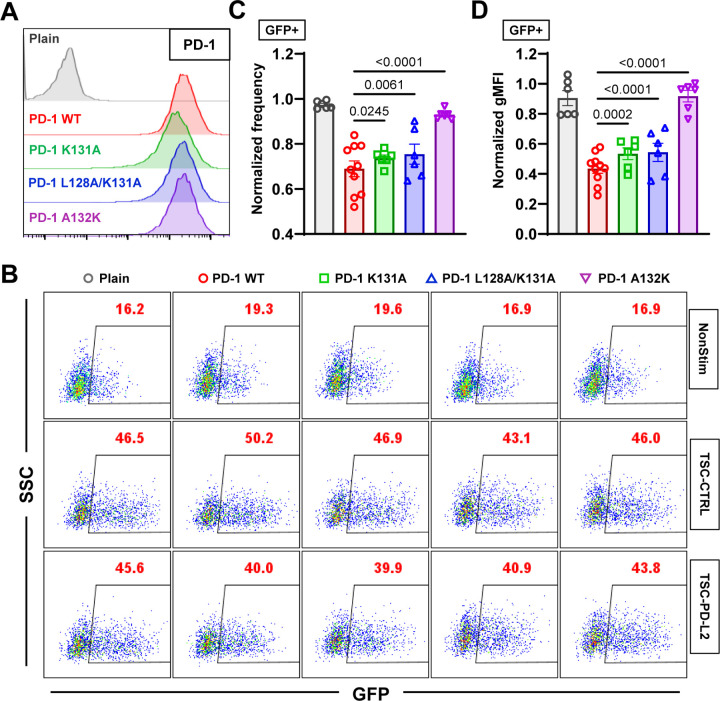
Immune checkpoint blockade targeting PD-1 shows great success in cancer therapy. However, the mechanism of how ligand binding initiates PD-1 signaling remains unclear. As prognosis markers of multiple cancers, soluble PD-L1 is found in patient sera and can bind PD-1, but fails to suppress T cell function. This and our previous observations that T cells exert endogenous forces on PD-1-PD-L2 bonds prompt the hypothesis that mechanical force might be critical to PD-1 triggering, which is missing in the soluble ligand case due to the lack of mechanical support afforded by surface-anchored ligand. Here we show that PD-1 function is eliminated or reduced when mechanical support on ligand is removed or dampened, respectively. Force spectroscopic analysis reveals that PD-1 forms catch bonds with both PD-Ligands <7 pN where force prolongs bond lifetime, but slip bonds >8 pN where force accelerates dissociation. Steered molecular dynamics finds PD-1-PD-L2 complex very sensitive to force due to the two molecules' "side-to-side" binding via β sheets. Pulling causes relative rotation and translation between the two molecules by stretching and aligning the complex along the force direction, yielding new atomic contacts not observed in the crystal structure. Compared to wild-type, PD-1 mutants targeting the force-induced new interactions maintain the same binding affinity but display lower rupture force, shorter bond lifetime, reduced tension, and most importantly, impaired capacity to suppress T cell activation. Our results uncover a mechanism for cells to probe the mechanical support of PD-1-PD-Ligand bonds using endogenous forces to regulate PD-1 triggering.
Figures
References
-
- Barber D. L. et al. Restoring function in exhausted CD8 T cells during chronic viral infection. Nature 439, 682–687 (2006). - PubMed
-
- Day C. L. et al. PD-1 expression on HIV-specific T cells is associated with T-cell exhaustion and disease progression. Nature 443, 350–354 (2006). - PubMed
-
- Jin H. T., Ahmed R. & Okazaki T. Role of PD-1 in regulating T-cell immunity. Current topics in microbiology and immunology 350, 17–37 (2011). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials