

The awakening of dormant neuronal precursors in the adult and aged brain
- PMID: 37649323
- PMCID: PMC10726842
- DOI: 10.1111/acel.13974
The awakening of dormant neuronal precursors in the adult and aged brain
Abstract
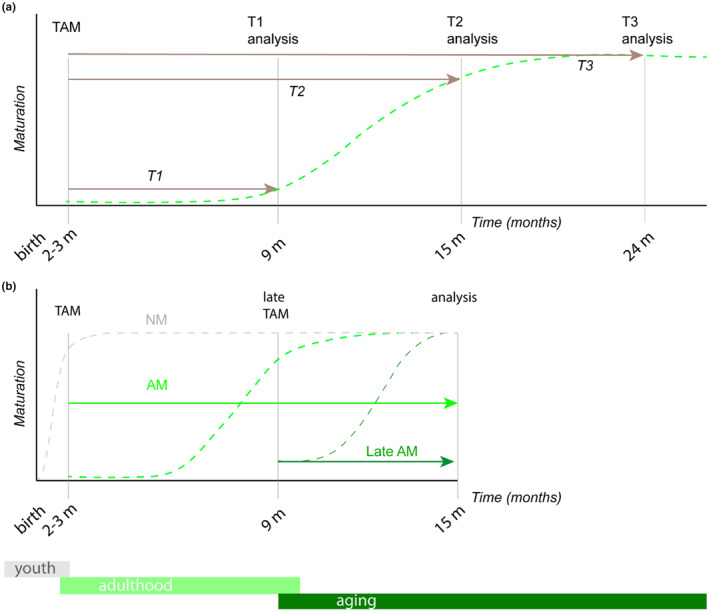
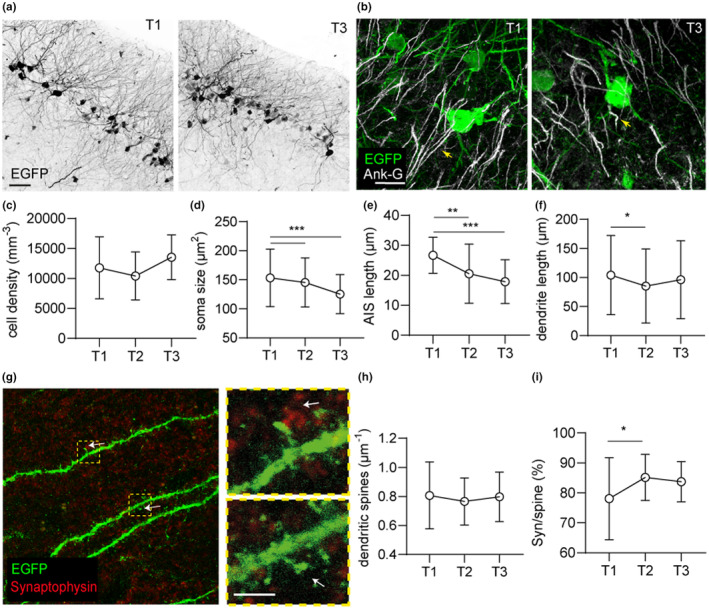
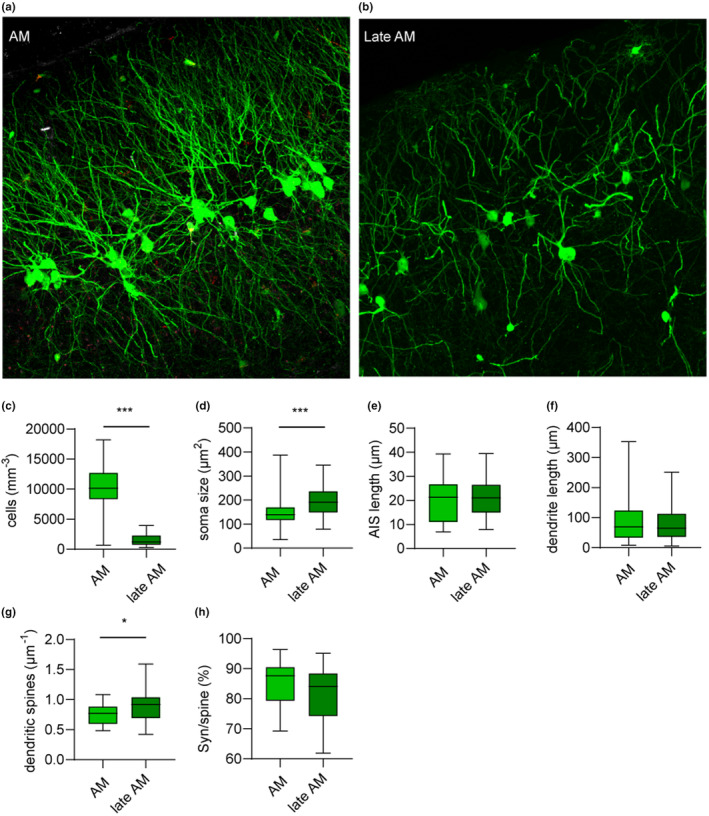
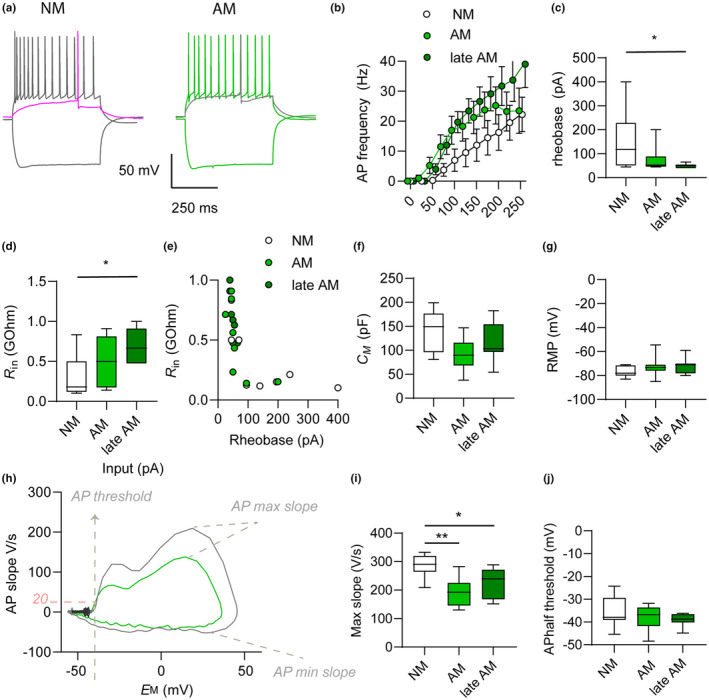
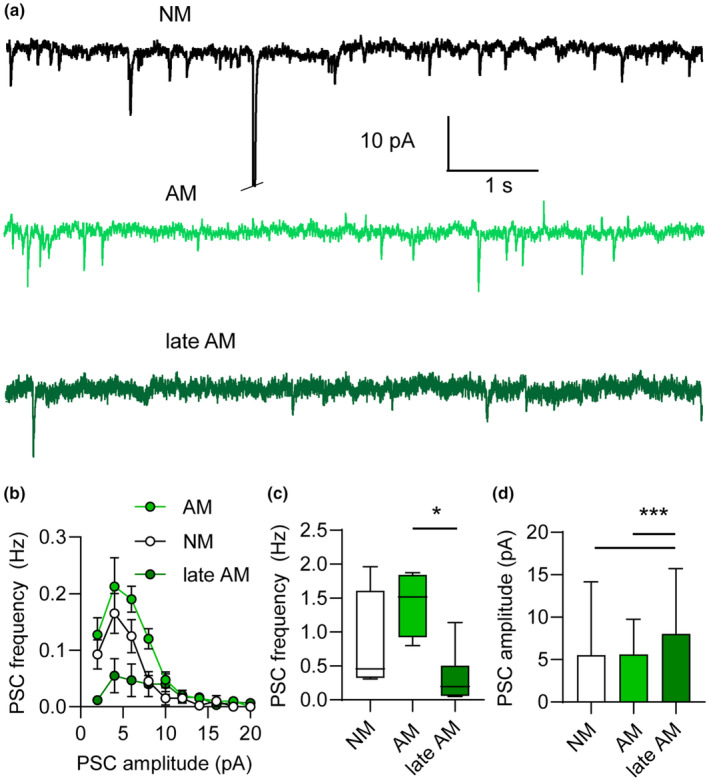
Beyond the canonical neurogenic niches, there are dormant neuronal precursors in several regions of the adult mammalian brain. Dormant precursors maintain persisting post-mitotic immaturity from birth to adulthood, followed by staggered awakening, in a process that is still largely unresolved. Strikingly, due to the slow rate of awakening, some precursors remain immature until old age, which led us to question whether their awakening and maturation are affected by aging. To this end, we studied the maturation of dormant precursors in transgenic mice (DCX-CreERT2 /flox-EGFP) in which immature precursors were labelled permanently in vivo at different ages. We found that dormant precursors are capable of awakening at young age, becoming adult-matured neurons (AM), as well as of awakening at old age, becoming late AM. Thus, protracted immaturity does not prevent late awakening and maturation. However, late AM diverged morphologically and functionally from AM. Moreover, AM were functionally most similar to neonatal-matured neurons (NM). Conversely, late AM were endowed with high intrinsic excitability and high input resistance, and received a smaller amount of spontaneous synaptic input, implying their relative immaturity. Thus, late AM awakening still occurs at advanced age, but the maturation process is slow.
Keywords: action potential; aged brain; axon initial segment; dormant precursor; doublecortin; neurogenesis; neuronal precursor; synapse.
© 2023 The Authors. Aging Cell published by Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Functional Integration of Neuronal Precursors in the Adult Murine Piriform Cortex.Cereb Cortex. 2020 Mar 14;30(3):1499-1515. doi: 10.1093/cercor/bhz181. Cereb Cortex. 2020. PMID: 31647533 Free PMC article.
-
Why Would the Brain Need Dormant Neuronal Precursors?Front Neurosci. 2022 Apr 8;16:877167. doi: 10.3389/fnins.2022.877167. eCollection 2022. Front Neurosci. 2022. PMID: 35464307 Free PMC article.
-
Cellular Plasticity in the Adult Murine Piriform Cortex: Continuous Maturation of Dormant Precursors Into Excitatory Neurons.Cereb Cortex. 2018 Jul 1;28(7):2610-2621. doi: 10.1093/cercor/bhy087. Cereb Cortex. 2018. PMID: 29688272 Free PMC article.
-
PSA Depletion Induces the Differentiation of Immature Neurons in the Piriform Cortex of Adult Mice.Int J Mol Sci. 2021 May 27;22(11):5733. doi: 10.3390/ijms22115733. Int J Mol Sci. 2021. PMID: 34072166 Free PMC article.
-
Consistency and Variation in Doublecortin and Ki67 Antigen Detection in the Brain Tissue of Different Mammals, including Humans.Int J Mol Sci. 2023 Jan 28;24(3):2514. doi: 10.3390/ijms24032514. Int J Mol Sci. 2023. PMID: 36768845 Free PMC article.
Cited by
-
Adult neurogenesis and "immature" neurons in mammals: an evolutionary trade-off in plasticity?Brain Struct Funct. 2024 Nov;229(8):1775-1793. doi: 10.1007/s00429-023-02717-9. Epub 2023 Oct 13. Brain Struct Funct. 2024. PMID: 37833544 Free PMC article. Review.
-
Phylogenetic variation of layer II cortical immature neurons in dog and horse confirms covariance with brain size and neocortical surface.Brain Struct Funct. 2025 Jul 7;230(6):115. doi: 10.1007/s00429-025-02981-x. Brain Struct Funct. 2025. PMID: 40622463 Free PMC article.
-
Multispecies characterization of immature neurons in the mammalian amygdala reveals their expansion in primates.PLoS Biol. 2025 Aug 14;23(8):e3003322. doi: 10.1371/journal.pbio.3003322. eCollection 2025 Aug. PLoS Biol. 2025. PMID: 40811712 Free PMC article.
-
Neuronal Subtypes and Connectivity of the Adult Mouse Paralaminar Amygdala.eNeuro. 2024 Jun 18;11(6):ENEURO.0119-24.2024. doi: 10.1523/ENEURO.0119-24.2024. Print 2024 Jun. eNeuro. 2024. PMID: 38811163 Free PMC article.
-
Neuron and Brain Maturation 2.0.Int J Mol Sci. 2023 Dec 4;24(23):17113. doi: 10.3390/ijms242317113. Int J Mol Sci. 2023. PMID: 38069434 Free PMC article.
References
-
- Abbott, L. C. , & Nigussie, F. (2020). Adult neurogenesis in the mammalian dentate gyrus. Journal of Veterinary Medicine Series C: Anatomia Histologia Embryologia, 49, 3–16. - PubMed
-
- Benedetti, B. , Dannehl, D. , König, R. , Coviello, S. , Kreutzer, C. , Zaunmair, P. , Jakubecova, D. , Weiger, T. M. , Aigner, L. , Nacher, J. , Engelhardt, M. , & Couillard‐Després, S. (2019). Functional integration of neuronal precursors in the adult murine piriform cortex. Cerebral Cortex, 30(3), 1499–1515. - PMC - PubMed
