

Developmental reprogramming of myometrial stem cells by endocrine disruptor linking to risk of uterine fibroids
- PMID: 37650943
- PMCID: PMC10471700
- DOI: 10.1007/s00018-023-04919-0
Developmental reprogramming of myometrial stem cells by endocrine disruptor linking to risk of uterine fibroids
Update in
-
Endocrine-disrupting chemicals and epigenetic reprogramming in developmental origin of uterine fibroids.Sci Prog. 2023 Oct-Dec;106(4):368504231215601. doi: 10.1177/00368504231215601. Sci Prog. 2023. PMID: 38189295 Free PMC article.
Abstract
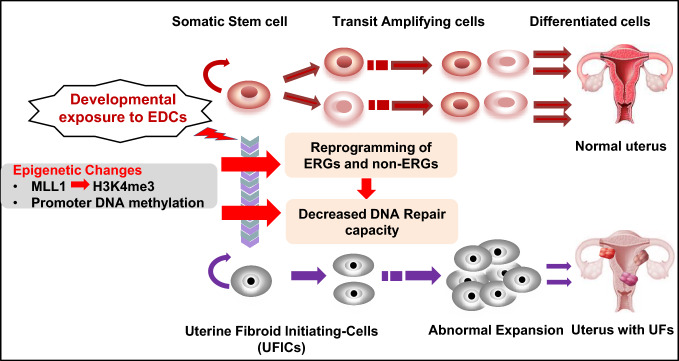
Background: The stage, when tissues and organs are growing, is very vulnerable to environmental influences, but it's not clear how exposure during this time causes changes to the epigenome and increases the risk of hormone-related illnesses like uterine fibroids (UFs).
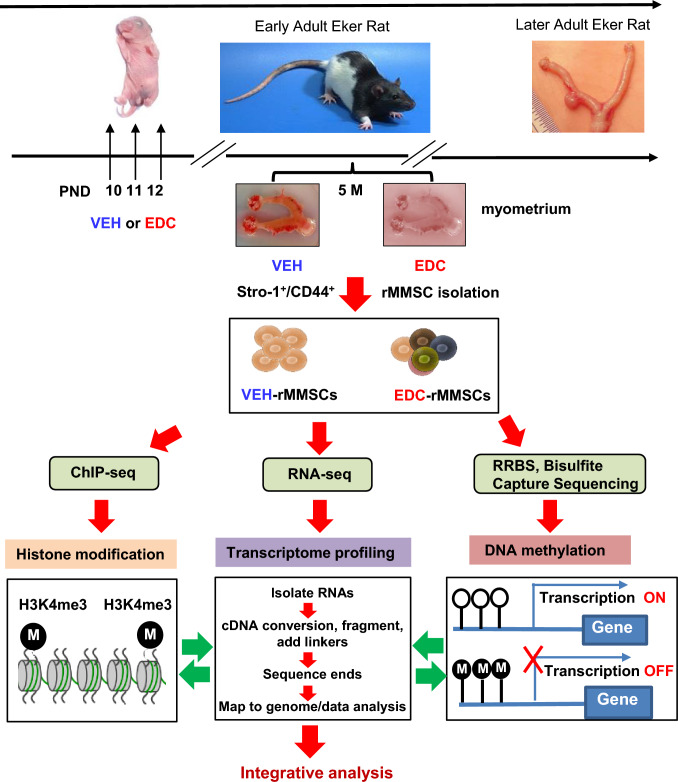
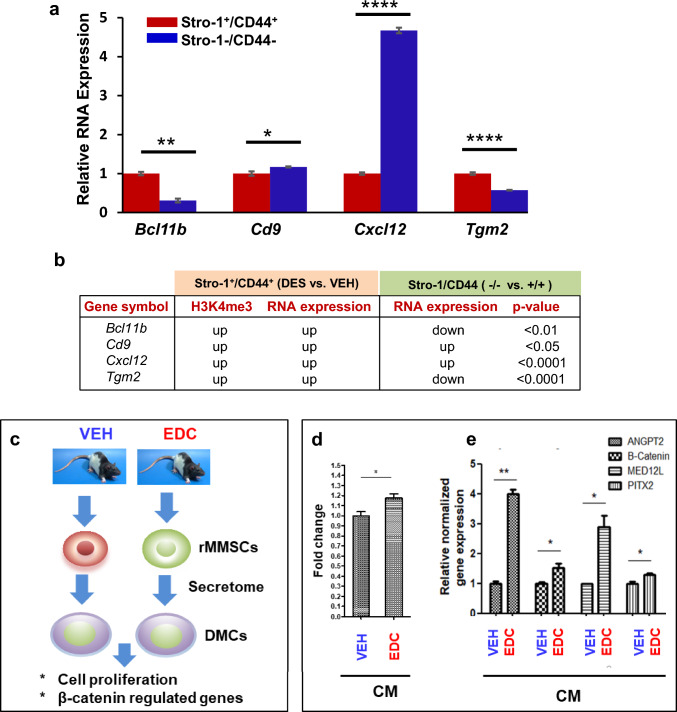
Methods: Developmental reprogramming of myometrial stem cells (MMSCs), the putative origin from which UFs originate, was investigated in vitro and in the Eker rat model by RNA-seq, ChIP-seq, RRBS, gain/loss of function analysis, and luciferase activity assays.
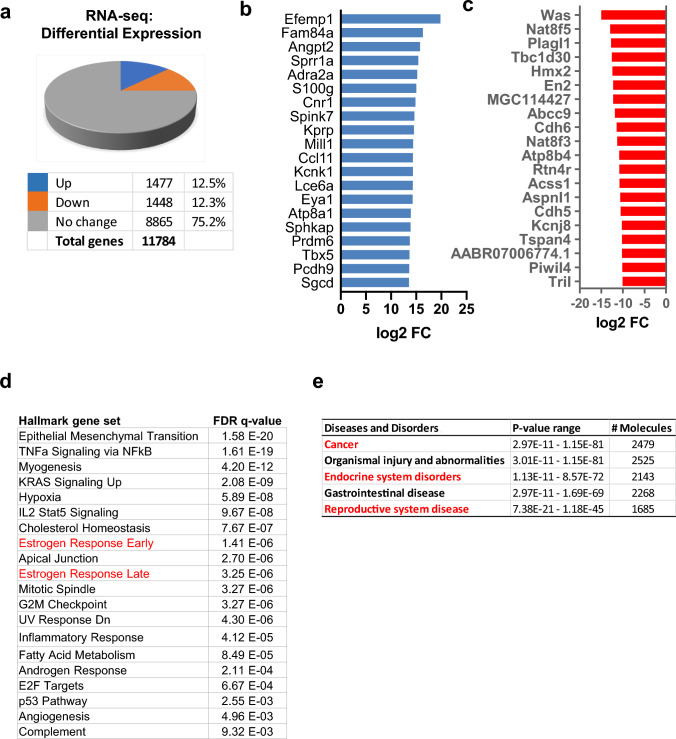
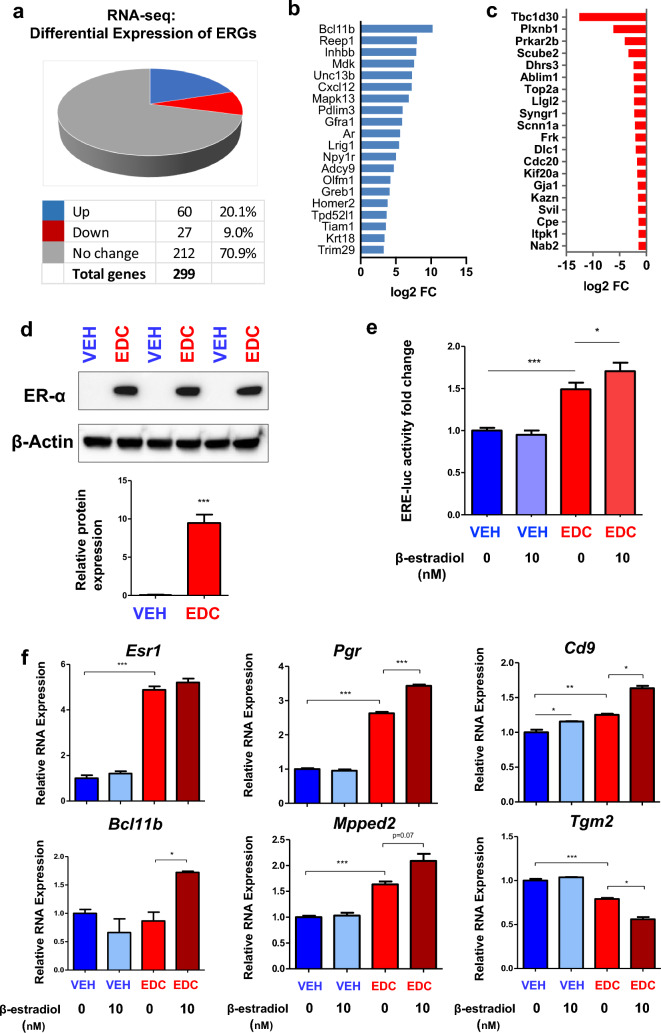
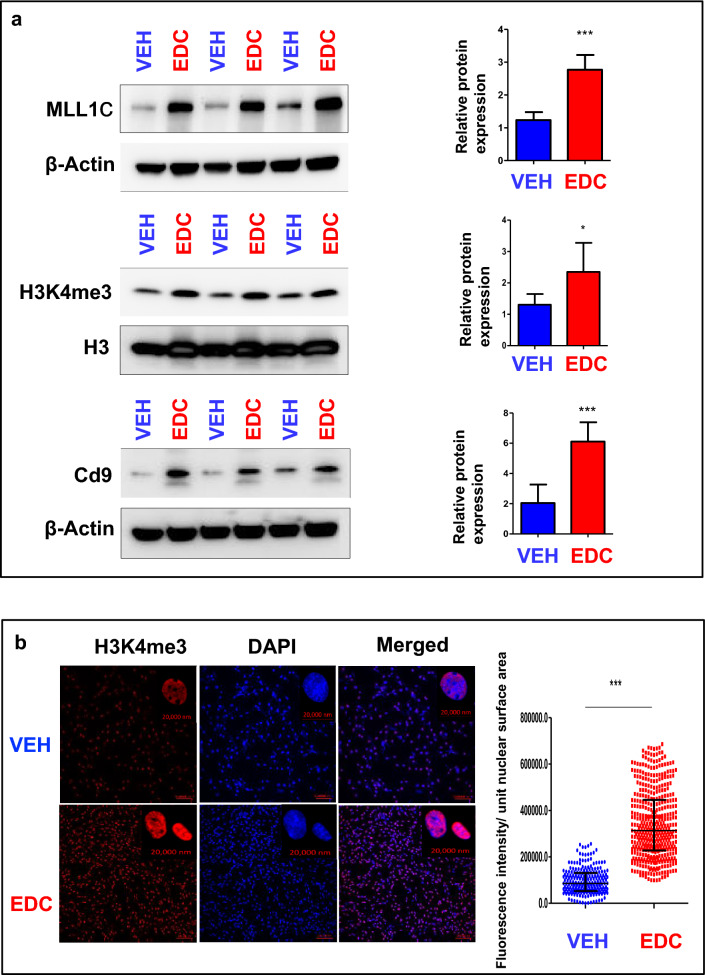
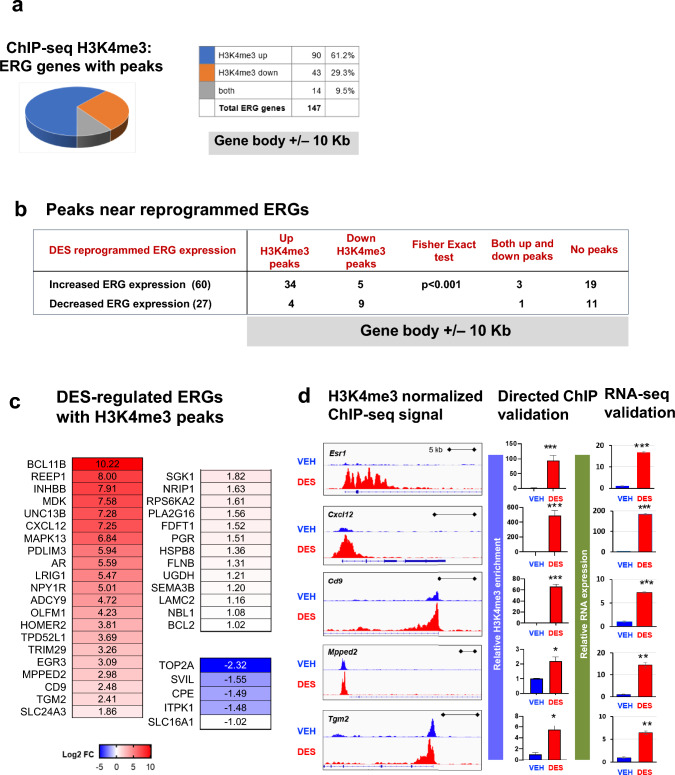
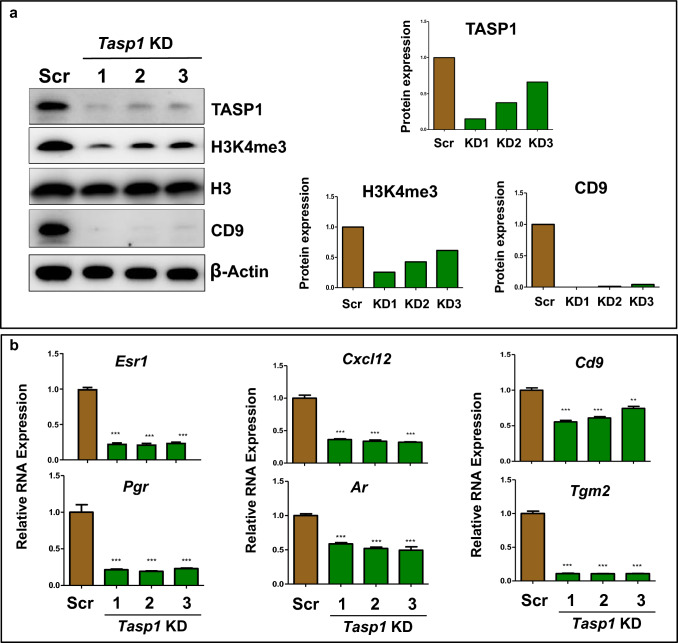
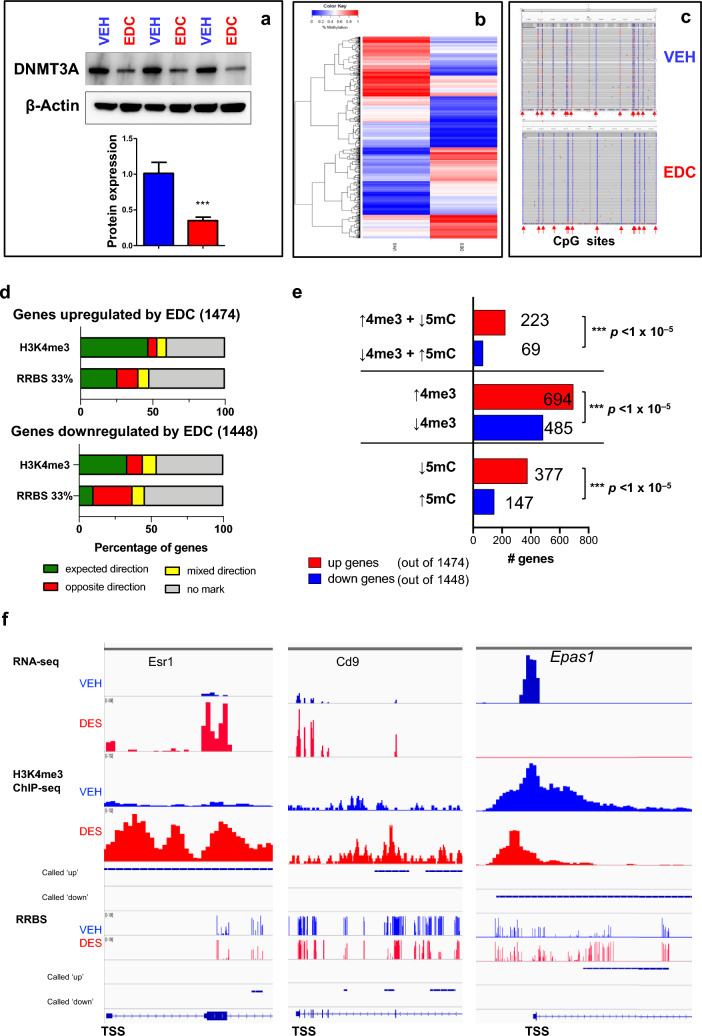
Results: When exposed to the endocrine-disrupting chemical (EDC) diethylstilbestrol during Eker rat development, MMSCs undergo a reprogramming of their estrogen-responsive transcriptome. The reprogrammed genes in MMSCs are known as estrogen-responsive genes (ERGs) and are activated by mixed lineage leukemia protein-1 (MLL1) and DNA hypo-methylation mechanisms. Additionally, we observed a notable elevation in the expression of ERGs in MMSCs from Eker rats exposed to natural steroids after developmental exposure to EDC, thereby augmenting estrogen activity.
Conclusion: Our studies identify epigenetic mechanisms of MLL1/DNA hypo-methylation-mediated MMSC reprogramming. EDC exposure epigenetically targets MMSCs and leads to persistent changes in the expression of a subset of ERGs, imparting a hormonal imprint on the ERGs, resulting in a "hyper-estrogenic" phenotype, and increasing the hormone-dependent risk of UFs.
Keywords: DNA hypo-methylation; Developmental reprogramming; Eker rat; Endocrine-disrupting chemicals; Epigenome; Estrogen signaling; Estrogen-responsive genes; Hormonal imprint; Hyper-estrogenic; Leiomyoma; MLL1 activation; Progenitor cells; TASP1.
© 2023. The Author(s).
Conflict of interest statement
None of the authors has a financial relationship with a commercial entity that has an interest in the subject of this manuscript.
Figures
References
-
- Bulun SE. Uterine fibroids. N Engl J Med. 2013;369(14):1344–1355. - PubMed
-
- Stewart EA, Laughlin-Tommaso SK, Catherino WH, Lalitkumar S, Gupta D, Vollenhoven B. Uterine fibroids. Nat Rev Dis Primers. 2016;2:16043. - PubMed
-
- Segars JH, Parrott EC, Nagel JD, Guo XC, Gao X, Birnbaum LS, Pinn VW, Dixon D. Proceedings from the Third National Institutes of Health International Congress on Advances in Uterine Leiomyoma Research: comprehensive review, conference summary and future recommendations. Hum Reprod Update. 2014;20(3):309–333. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
