

A lipid-associated macrophage lineage rewires the spatial landscape of adipose tissue in early obesity
- PMID: 37651193
- PMCID: PMC10619435
- DOI: 10.1172/jci.insight.171701
A lipid-associated macrophage lineage rewires the spatial landscape of adipose tissue in early obesity
Abstract
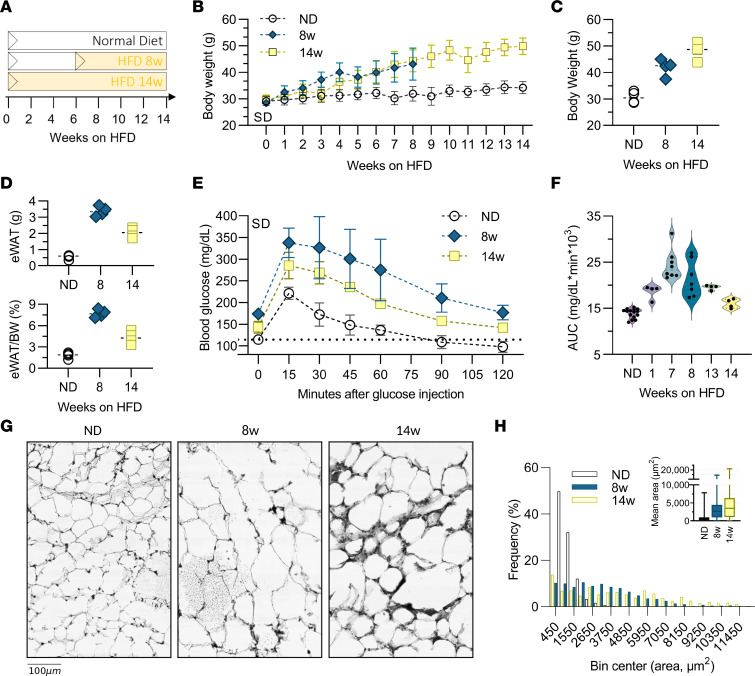
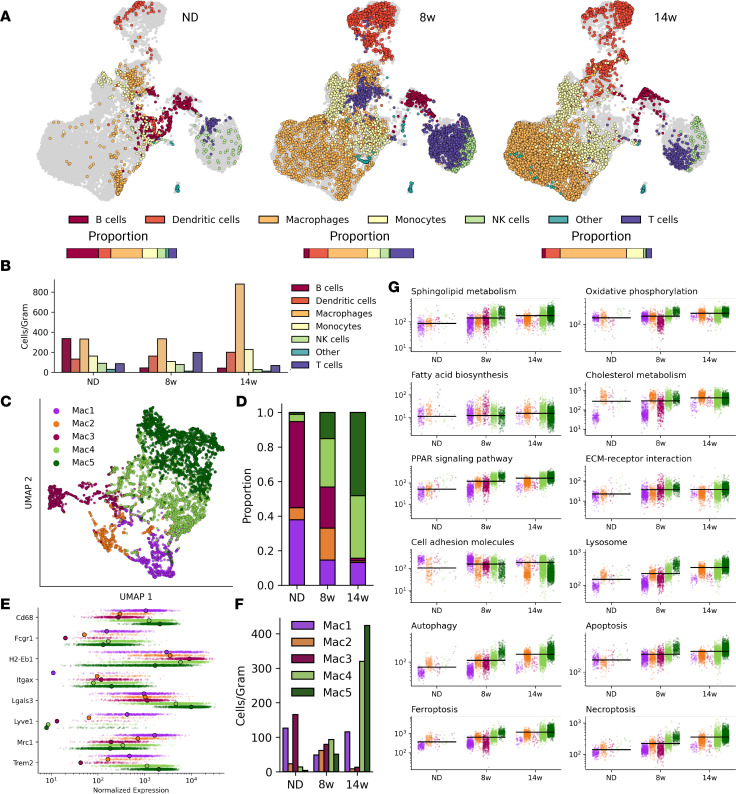
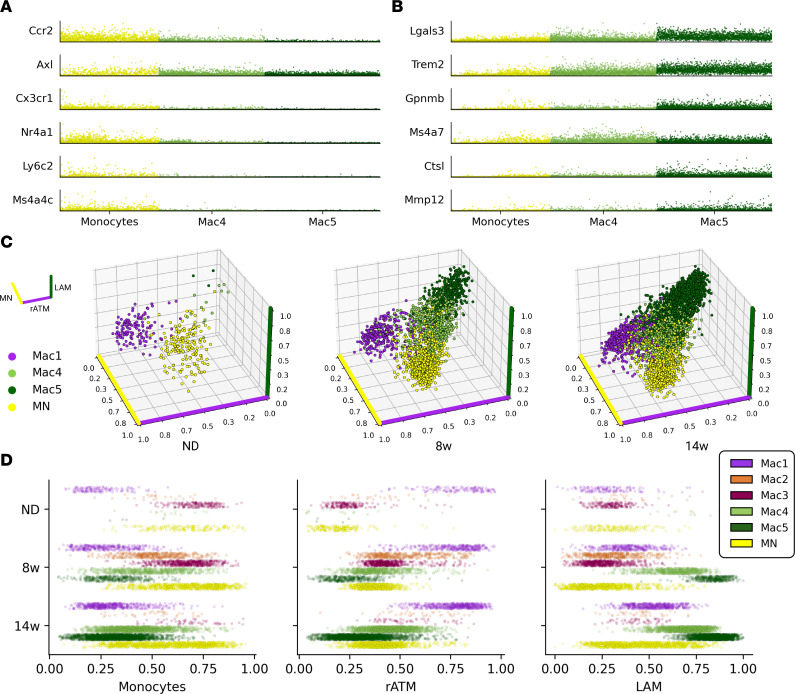
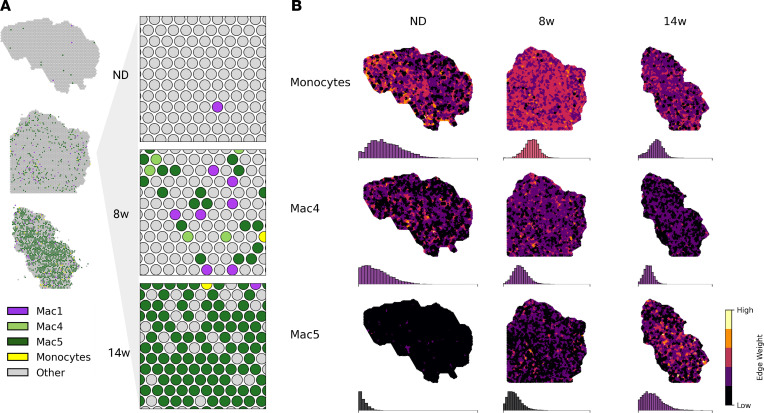
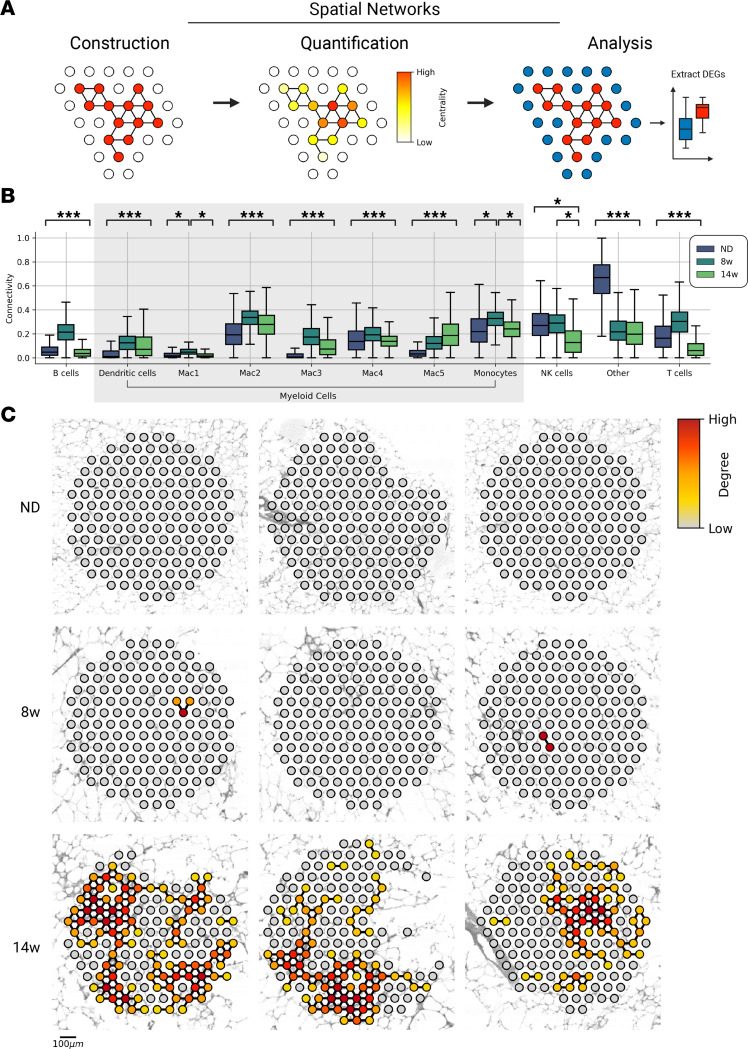
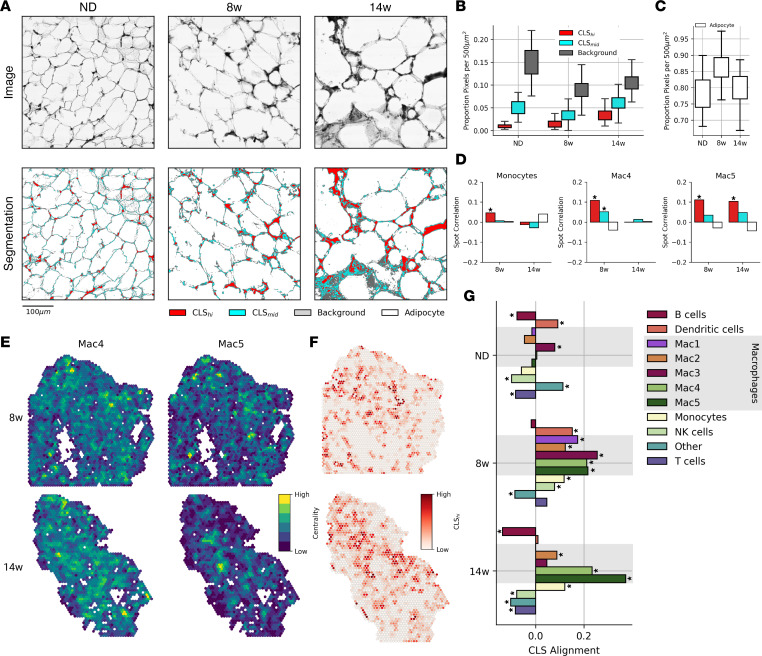
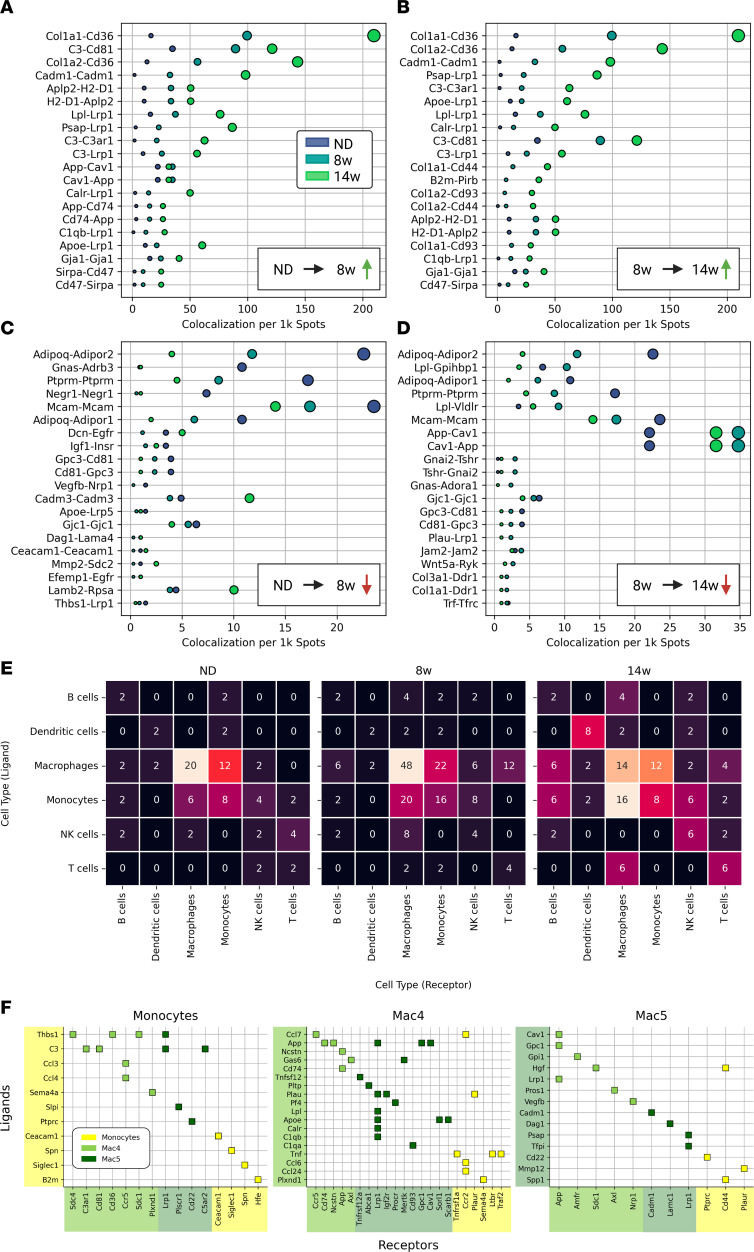
Adipose tissue macrophage (ATM) infiltration is associated with adipose tissue dysfunction and insulin resistance in mice and humans. Recent single-cell data highlight increased ATM heterogeneity in obesity but do not provide a spatial context for ATM phenotype dynamics. We integrated single-cell RNA-Seq, spatial transcriptomics, and imaging of murine adipose tissue in a time course study of diet-induced obesity. Overall, proinflammatory immune cells were predominant in early obesity, whereas nonresident antiinflammatory ATMs predominated in chronic obesity. A subset of these antiinflammatory ATMs were transcriptomically intermediate between monocytes and mature lipid-associated macrophages (LAMs) and were consistent with a LAM precursor (pre-LAM). Pre-LAMs were spatially associated with early obesity crown-like structures (CLSs), which indicate adipose tissue dysfunction. Spatial data showed colocalization of ligand-receptor transcripts related to lipid signaling among monocytes, pre-LAMs, and LAMs, including Apoe, Lrp1, Lpl, and App. Pre-LAM expression of these ligands in early obesity suggested signaling to LAMs in the CLS microenvironment. Our results refine understanding of ATM diversity and provide insight into the dynamics of the LAM lineage during development of metabolic disease.
Keywords: Adipose tissue; Macrophages; Metabolism; Obesity.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
