

Inhibition of α-hemolysin activity of Staphylococcus aureus by theaflavin 3,3'-digallate
- PMID: 37651426
- PMCID: PMC10470925
- DOI: 10.1371/journal.pone.0290904
Inhibition of α-hemolysin activity of Staphylococcus aureus by theaflavin 3,3'-digallate
Abstract
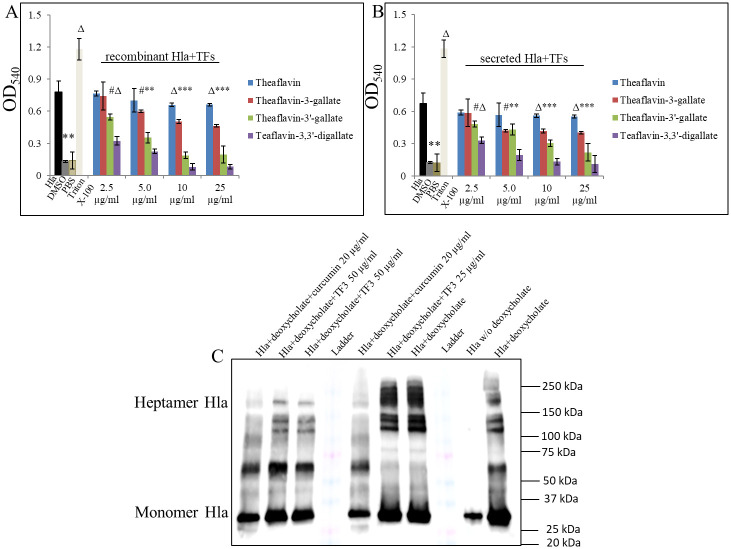
The ongoing rise in antibiotic resistance, and a waning of the introduction of new antibiotics, has resulted in limited treatment options for bacterial infections, including these caused by methicillin-resistant Staphylococcus aureus, leaving the world in a post-antibiotic era. Here, we set out to examine mechanisms by which theaflavin 3,3'-digallate (TF3) might act as an anti-hemolytic compound. In the presented study, we found that TF3 has weak bacteriostatic and bactericidal effects on Staphylococcus aureus, and strong inhibitory effect towards the hemolytic activity of its α-hemolysin (Hla) including its production and secretion. A supportive SPR assay reinforced these results and further revealed binding of TF3 to Hla with KD = 4.57×10-5 M. Interestingly, TF3 was also able to protect human primary keratinocytes from Hla-induced cell death, being at the same time non-toxic for them. Further analysis of TF3 properties revealed that TF3 blocked Hla-prompting immune reaction by inhibiting production and secretion of IL1β, IL6, and TNFα in vitro and in vivo, through affecting NFκB activity. Additionally, we observed that TF3 also markedly attenuated S. aureus-induced barrier disruption, by inhibiting Hla-triggered E-cadherin and ZO-1 impairment. Overall, by blocking activity of Hla, TF3 subsequently subdued the inflammation and protected the epithelial barrier, which is considered as beneficial to relieving skin injury.
Copyright: © 2023 Goc et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
