

Tumor-associated monocytes promote mesenchymal transformation through EGFR signaling in glioma
- PMID: 37652019
- PMCID: PMC10518634
- DOI: 10.1016/j.xcrm.2023.101177
Tumor-associated monocytes promote mesenchymal transformation through EGFR signaling in glioma
Abstract
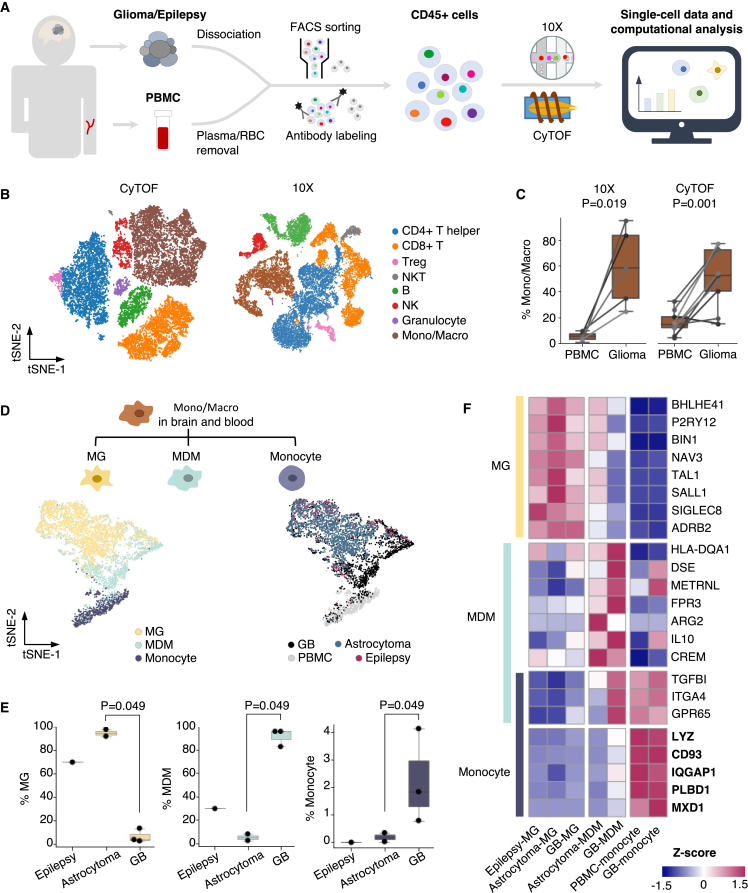
The role of brain immune compartments in glioma evolution remains elusive. We profile immune cells in glioma microenvironment and the matched peripheral blood from 11 patients. Glioblastoma exhibits specific infiltration of blood-originated monocytes expressing epidermal growth factor receptor (EGFR) ligands EREG and AREG, coined as tumor-associated monocytes (TAMo). TAMo infiltration is mutually exclusive with EGFR alterations (p = 0.019), while co-occurring with mesenchymal subtype (p = 4.7 × 10-7) and marking worse prognosis (p = 0.004 and 0.032 in two cohorts). Evolutionary analysis of initial-recurrent glioma pairs and single-cell study of a multi-centric glioblastoma reveal association between elevated TAMo and glioma mesenchymal transformation. Further analyses identify FOSL2 as a TAMo master regulator and demonstrates that FOSL2-EREG/AREG-EGFR signaling axis promotes glioma invasion in vitro. Collectively, we identify TAMo in tumor microenvironment and reveal its driving role in activating EGFR signaling to shape glioma evolution.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Similar articles
-
Nuclear FABP7 immunoreactivity is preferentially expressed in infiltrative glioma and is associated with poor prognosis in EGFR-overexpressing glioblastoma.BMC Cancer. 2006 Apr 19;6:97. doi: 10.1186/1471-2407-6-97. BMC Cancer. 2006. PMID: 16623952 Free PMC article.
-
A comprehensive study of the association between the EGFR and ERBB2 genes and glioma risk.Acta Oncol. 2010 Aug;49(6):767-75. doi: 10.3109/0284186X.2010.480980. Acta Oncol. 2010. PMID: 20446891
-
EGFR mutation: novel prognostic factor associated with immune infiltration in lower-grade glioma; an exploratory study.BMC Cancer. 2019 Dec 4;19(1):1184. doi: 10.1186/s12885-019-6384-8. BMC Cancer. 2019. PMID: 31801484 Free PMC article.
-
Updated Insights on EGFR Signaling Pathways in Glioma.Int J Mol Sci. 2021 Jan 8;22(2):587. doi: 10.3390/ijms22020587. Int J Mol Sci. 2021. PMID: 33435537 Free PMC article. Review.
-
An Overview of EGFR Mechanisms and Their Implications in Targeted Therapies for Glioblastoma.Int J Mol Sci. 2023 Jul 5;24(13):11110. doi: 10.3390/ijms241311110. Int J Mol Sci. 2023. PMID: 37446288 Free PMC article. Review.
Cited by
-
Single-cell RNA sequencing identifies a subtype of FN1 + tumor-associated macrophages associated with glioma recurrence and as a biomarker for immunotherapy.Biomark Res. 2024 Oct 7;12(1):114. doi: 10.1186/s40364-024-00662-1. Biomark Res. 2024. PMID: 39375795 Free PMC article.
-
Sex differences in the molecular profile of adult diffuse glioma are shaped by IDH status and tumor microenvironment.Neuro Oncol. 2025 Feb 10;27(2):430-444. doi: 10.1093/neuonc/noae207. Neuro Oncol. 2025. PMID: 39367624
-
Integrating Multiplex Immunohistochemistry and Machine Learning for Glioma Subtyping and Prognosis Prediction.MedComm (2020). 2025 Apr 22;6(5):e70138. doi: 10.1002/mco2.70138. eCollection 2025 May. MedComm (2020). 2025. PMID: 40264576 Free PMC article.
-
A Spatial Multi-Omic Framework Identifies Gliomas Permissive to TIL Expansion.Res Sq [Preprint]. 2025 Apr 25:rs.3.rs-6314842. doi: 10.21203/rs.3.rs-6314842/v1. Res Sq. 2025. PMID: 40313763 Free PMC article. Preprint.
-
Investigation of MANF regulation of glioma stemness via STAT3/TGF-β/SMAD4/p38 pathway based on pan-cancer analysis.Transl Oncol. 2025 Oct;60:102497. doi: 10.1016/j.tranon.2025.102497. Epub 2025 Aug 9. Transl Oncol. 2025. PMID: 40789259 Free PMC article.
References
-
- Cloughesy T.F., Mochizuki A.Y., Orpilla J.R., Hugo W., Lee A.H., Davidson T.B., Wang A.C., Ellingson B.M., Rytlewski J.A., Sanders C.M., et al. Neoadjuvant anti-PD-1 immunotherapy promotes a survival benefit with intratumoral and systemic immune responses in recurrent glioblastoma. Nat. Med. 2019;25:477–486. doi: 10.1038/s41591-018-0337-7. - DOI - PMC - PubMed
-
- De Groot J., Penas-Prado M., Alfaro-Munoz K., Hunter K., Pei B.L., O’Brien B., Weathers S.P., Loghin M., Kamiya Matsouka C., Yung W.K.A., et al. Window-of-opportunity clinical trial of pembrolizumab in patients with recurrent glioblastoma reveals predominance of immune-suppressive macrophages. Neuro Oncol. 2020;22:539–549. doi: 10.1093/neuonc/noz185. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
