

Birds multiplex spectral and temporal visual information via retinal On- and Off-channels
- PMID: 37652912
- PMCID: PMC10471707
- DOI: 10.1038/s41467-023-41032-z
Birds multiplex spectral and temporal visual information via retinal On- and Off-channels
Abstract
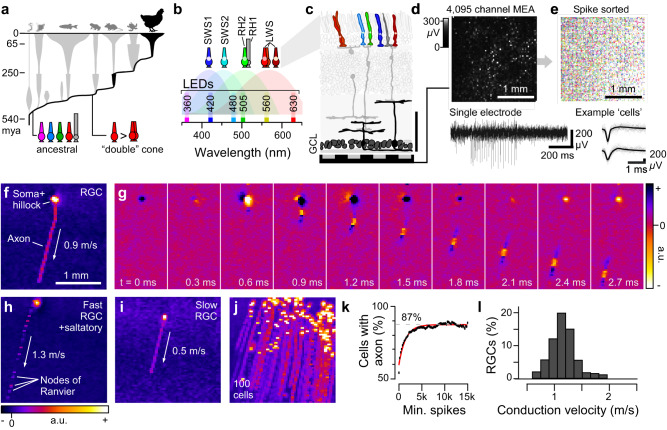
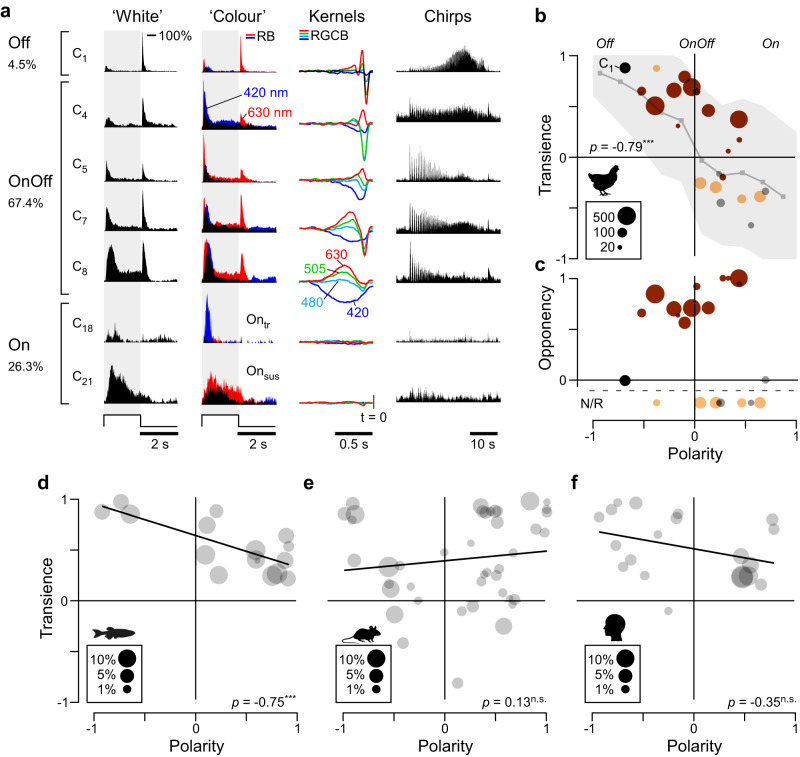
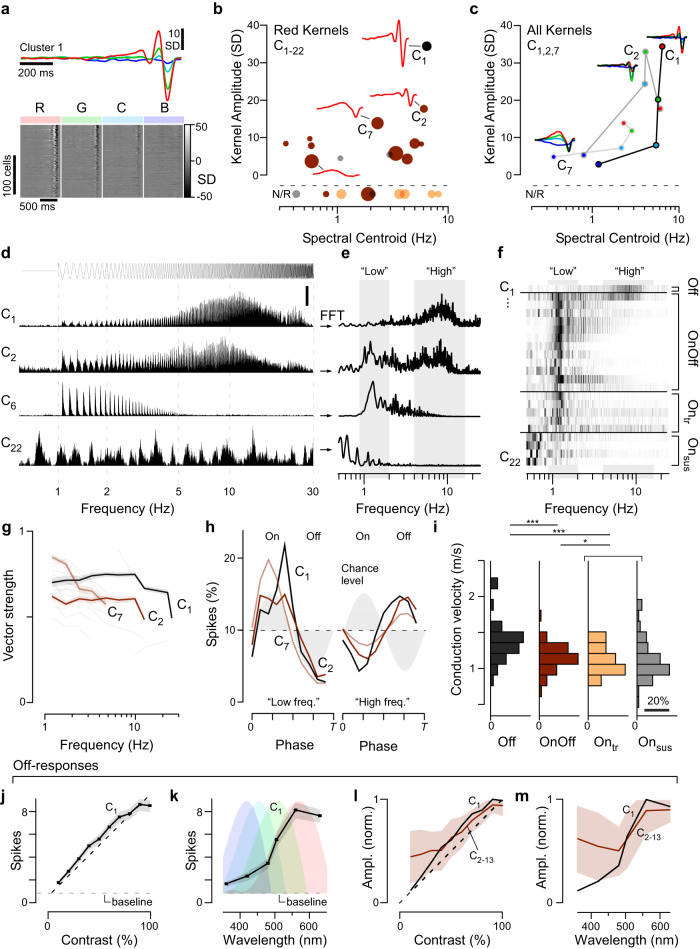
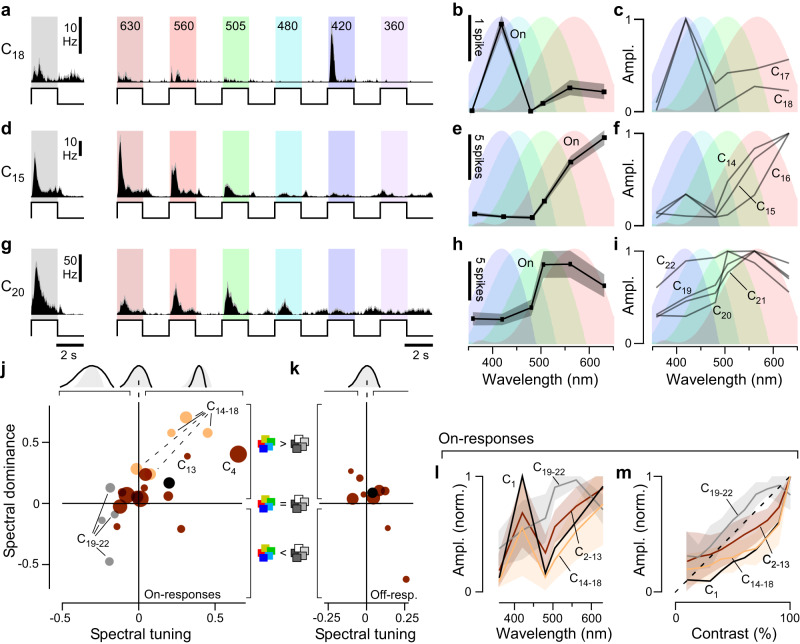
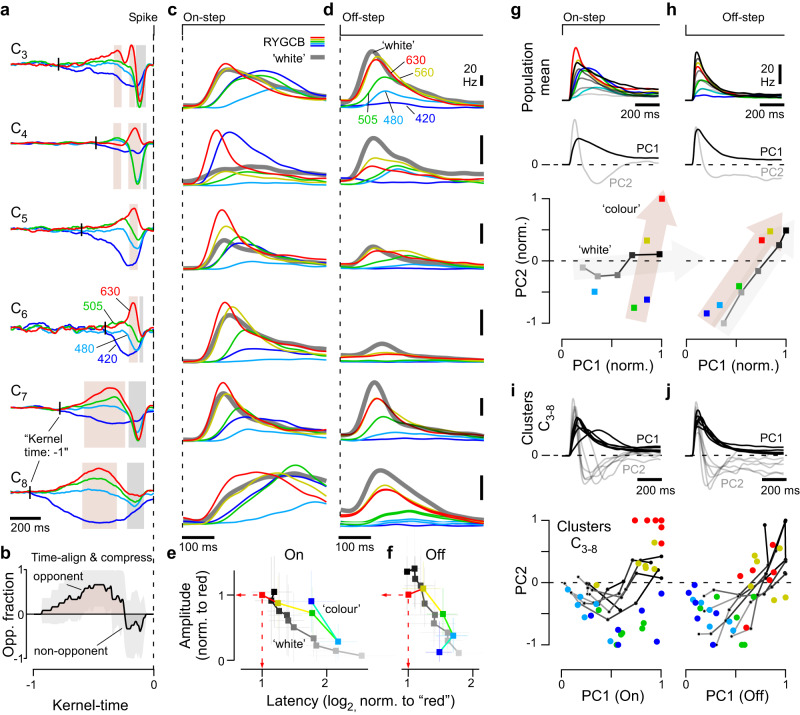
In vertebrate vision, early retinal circuits divide incoming visual information into functionally opposite elementary signals: On and Off, transient and sustained, chromatic and achromatic. Together these signals can yield an efficient representation of the scene for transmission to the brain via the optic nerve. However, this long-standing interpretation of retinal function is based on mammals, and it is unclear whether this functional arrangement is common to all vertebrates. Here we show that male poultry chicks use a fundamentally different strategy to communicate information from the eye to the brain. Rather than using functionally opposite pairs of retinal output channels, chicks encode the polarity, timing, and spectral composition of visual stimuli in a highly correlated manner: fast achromatic information is encoded by Off-circuits, and slow chromatic information overwhelmingly by On-circuits. Moreover, most retinal output channels combine On- and Off-circuits to simultaneously encode, or multiplex, both achromatic and chromatic information. Our results from birds conform to evidence from fish, amphibians, and reptiles which retain the full ancestral complement of four spectral types of cone photoreceptors.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
The Retinal Basis of Vertebrate Color Vision.Annu Rev Vis Sci. 2019 Sep 15;5:177-200. doi: 10.1146/annurev-vision-091718-014926. Epub 2019 Jun 21. Annu Rev Vis Sci. 2019. PMID: 31226010 Review.
-
Robust cone-mediated signaling persists late into rod photoreceptor degeneration.Elife. 2022 Aug 30;11:e80271. doi: 10.7554/eLife.80271. Elife. 2022. PMID: 36040015 Free PMC article.
-
S cones: Evolution, retinal distribution, development, and spectral sensitivity.Vis Neurosci. 2014 Mar;31(2):115-38. doi: 10.1017/S0952523813000242. Epub 2013 Jul 29. Vis Neurosci. 2014. PMID: 23895771 Review.
-
Oblique color vision in an open-habitat bird: spectral sensitivity, photoreceptor distribution and behavioral implications.J Exp Biol. 2012 Oct 1;215(Pt 19):3442-52. doi: 10.1242/jeb.073957. J Exp Biol. 2012. PMID: 22956248
-
From water to land: Evolution of photoreceptor circuits for vision in air.PLoS Biol. 2024 Jan 22;22(1):e3002422. doi: 10.1371/journal.pbio.3002422. eCollection 2024 Jan. PLoS Biol. 2024. PMID: 38252616 Free PMC article.
Cited by
-
Asymmetric distribution of color-opponent response types across mouse visual cortex supports superior color vision in the sky.Elife. 2024 Sep 5;12:RP89996. doi: 10.7554/eLife.89996. Elife. 2024. PMID: 39234821 Free PMC article.
-
Morphology and connectivity of retinal horizontal cells in two avian species.Front Cell Neurosci. 2025 Mar 4;19:1558605. doi: 10.3389/fncel.2025.1558605. eCollection 2025. Front Cell Neurosci. 2025. PMID: 40103750 Free PMC article.
-
Ancestral photoreceptor diversity as the basis of visual behaviour.Nat Ecol Evol. 2024 Mar;8(3):374-386. doi: 10.1038/s41559-023-02291-7. Epub 2024 Jan 22. Nat Ecol Evol. 2024. PMID: 38253752 Review.
-
A standardized nomenclature for the rods and cones of the vertebrate retina.PLoS Biol. 2025 May 7;23(5):e3003157. doi: 10.1371/journal.pbio.3003157. eCollection 2025 May. PLoS Biol. 2025. PMID: 40333813 Free PMC article.
-
Long-term, high-resolution in vivo calcium imaging in pigeons.Cell Rep Methods. 2024 Feb 26;4(2):100711. doi: 10.1016/j.crmeth.2024.100711. Epub 2024 Feb 20. Cell Rep Methods. 2024. PMID: 38382523 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
