

The contribution of muscle spindles to position sense measured with three different methods
- PMID: 37653105
- PMCID: PMC10520194
- DOI: 10.1007/s00221-023-06689-4
The contribution of muscle spindles to position sense measured with three different methods
Abstract
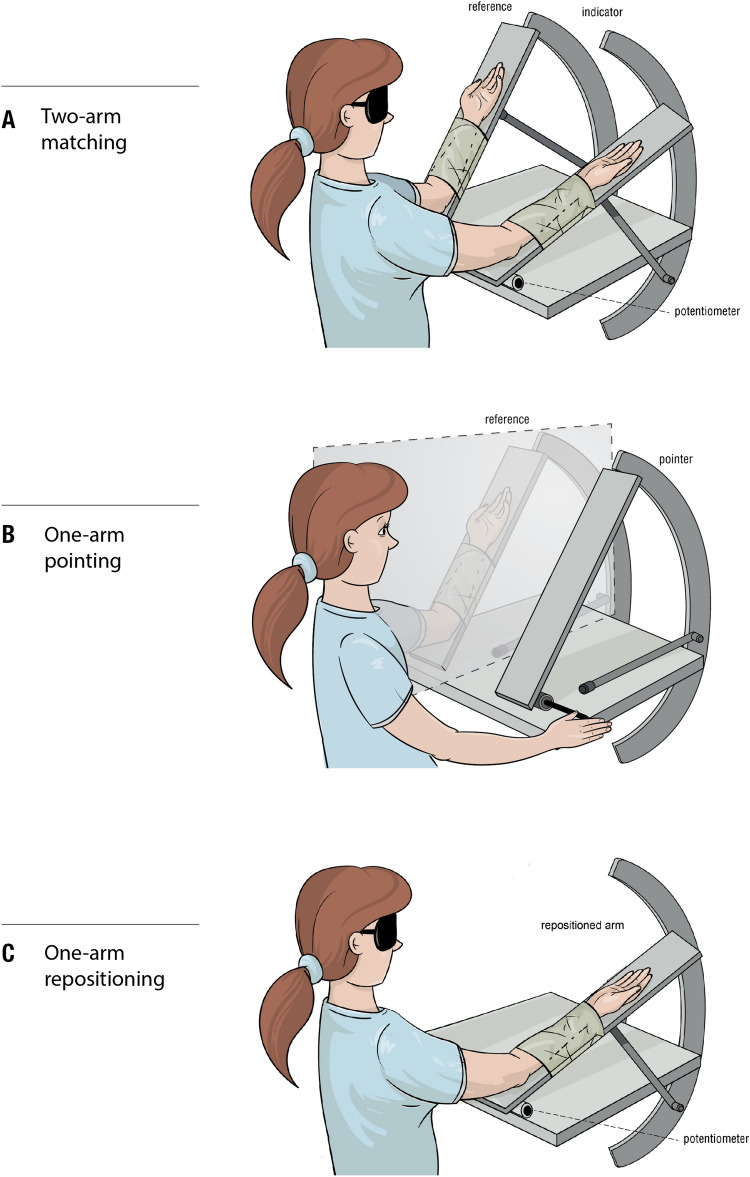
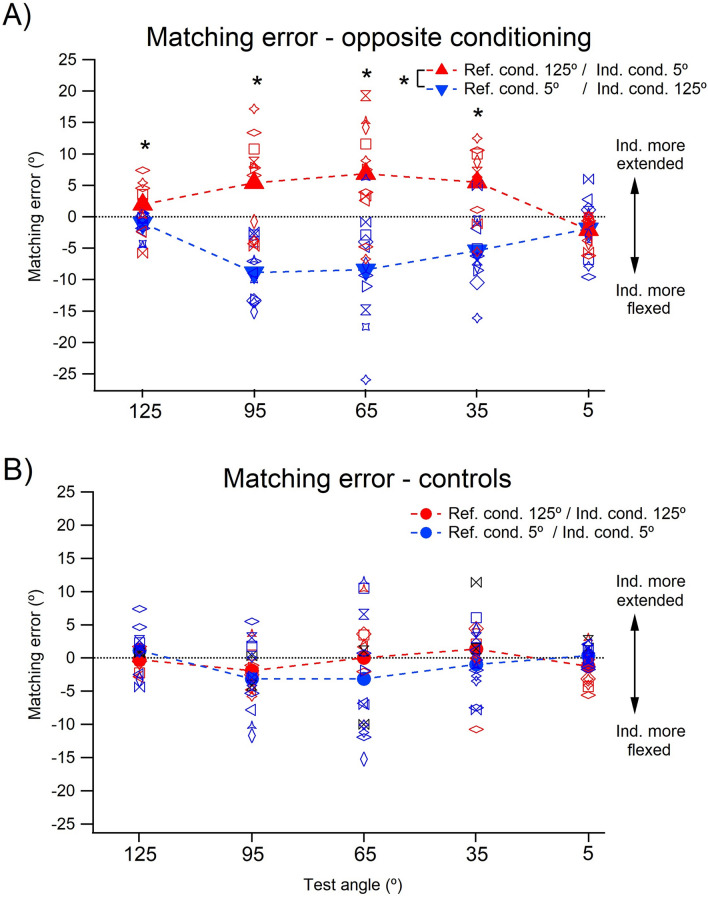
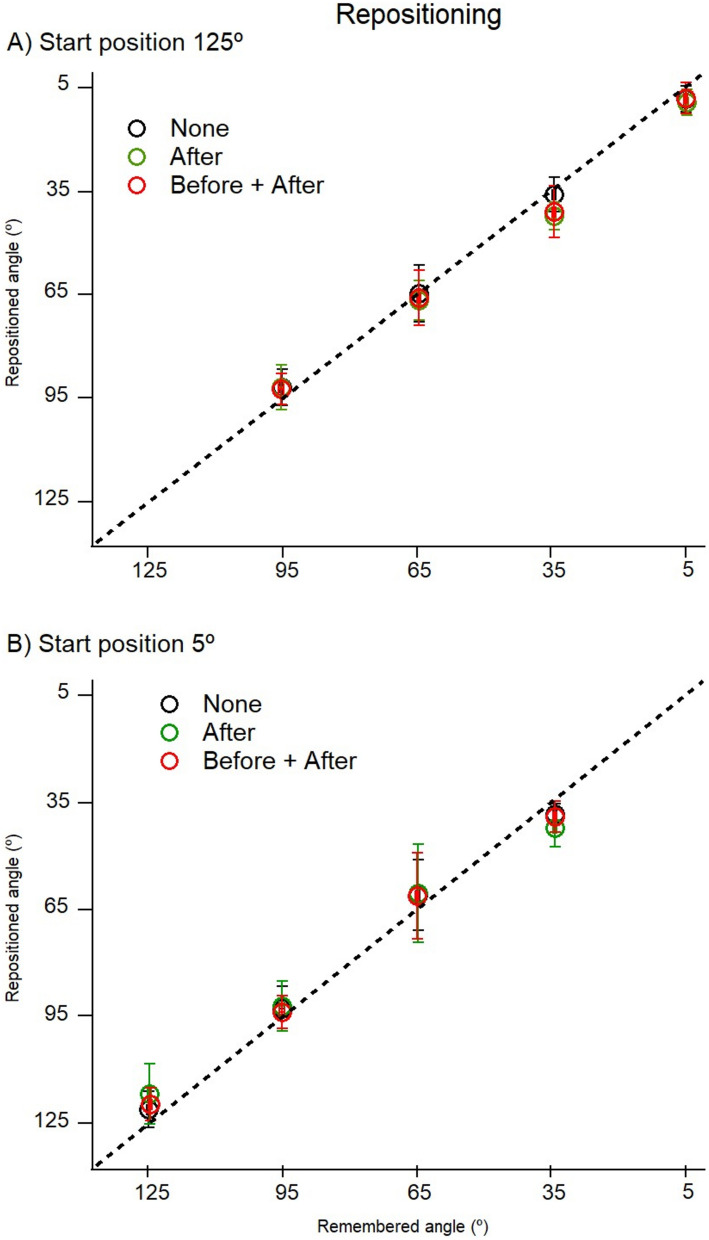
The sense of limb position is important, because it is believed to contribute to our sense of self-awareness. Muscle spindles, including both primary and secondary endings of spindles, are thought to be the principal position sensors. Passive spindles possess a property called thixotropy which allows their sensitivity to be manipulated. Here, thixotropic patterns of position errors have been studied with three commonly used methods of measurement of position sense. The patterns of errors have been used as indicators of the influence exerted by muscle spindles on a measured value of position sense. In two-arm matching, the blindfolded participant indicates the location of one arm by placement of the other. In one-arm pointing, the participant points to the perceived position of their other, hidden arm. In repositioning, one of the blindfolded participant's arms is placed at a chosen angle and they are asked to remember its position and then, after a delay, reproduce the position. The three methods were studied over the full range of elbow angles between 5° (elbow extension) and 125° (elbow flexion). Different outcomes were achieved with each method; in two-arm matching, position errors were symmetrical about zero and thixotropic influences were large, while in one-arm pointing, errors were biased towards extension. In repositioning, thixotropic effects were small. We conclude that each of the methods of measuring position sense comprises different mixes of peripheral and central influences. This will have to be taken into consideration by the clinician diagnosing disturbances in position sense.
Keywords: Muscle spindle; Proprioception; Self-awareness; Thixotropy.
© 2023. The Author(s).
Conflict of interest statement
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Figures



Similar articles
-
Position sense at the human elbow joint measured by arm matching or pointing.Exp Brain Res. 2016 Oct;234(10):2787-98. doi: 10.1007/s00221-016-4680-y. Epub 2016 May 21. Exp Brain Res. 2016. PMID: 27209194
-
Position sense at the human forearm over a range of elbow angles.Exp Brain Res. 2021 Feb;239(2):675-686. doi: 10.1007/s00221-020-05999-1. Epub 2021 Jan 3. Exp Brain Res. 2021. PMID: 33388906
-
Muscle thixotropy as a tool in the study of proprioception.Exp Brain Res. 2014 Nov;232(11):3397-412. doi: 10.1007/s00221-014-4088-5. Epub 2014 Sep 9. Exp Brain Res. 2014. PMID: 25200179 Review.
-
A reassessment of the role of joint receptors in human position sense.Exp Brain Res. 2023 Apr;241(4):943-949. doi: 10.1007/s00221-023-06582-0. Epub 2023 Mar 3. Exp Brain Res. 2023. PMID: 36869268 Free PMC article. Review.
-
The sensory origins of human position sense.J Physiol. 2016 Feb 15;594(4):1037-49. doi: 10.1113/JP271498. Epub 2016 Jan 19. J Physiol. 2016. PMID: 26537335 Free PMC article.
Cited by
-
How Multifunctioning Joints Produce Highly Agile Limbs in Animals with Lessons for Robotics.Biomimetics (Basel). 2024 Sep 3;9(9):529. doi: 10.3390/biomimetics9090529. Biomimetics (Basel). 2024. PMID: 39329551 Free PMC article. Review.
-
Effects of attentional focus on spatial localization of distal body parts and touch in two-arm position matching.Exp Brain Res. 2024 Dec 19;243(1):27. doi: 10.1007/s00221-024-06976-8. Exp Brain Res. 2024. PMID: 39699636
-
Comparison of joint position sense measured by inertial sensors embedded in portable digital devices with different masses.Front Neurosci. 2025 May 13;19:1561241. doi: 10.3389/fnins.2025.1561241. eCollection 2025. Front Neurosci. 2025. PMID: 40433496 Free PMC article.
-
Proprioception and its relationship with range of motion in hypermobile and normal mobile children.Exp Brain Res. 2024 Dec;242(12):2727-2735. doi: 10.1007/s00221-024-06937-1. Epub 2024 Oct 8. Exp Brain Res. 2024. PMID: 39377918 Free PMC article.
-
Normative values of spinal and peripheral proprioception in position sense among healthy adolescents and young adults.Sci Rep. 2024 Dec 30;14(1):31722. doi: 10.1038/s41598-024-82100-8. Sci Rep. 2024. PMID: 39738272 Free PMC article.
References
MeSH terms
LinkOut - more resources
Full Text Sources
