

The seasons within: a theoretical perspective on photoperiodic entrainment and encoding
- PMID: 37659985
- PMCID: PMC11226496
- DOI: 10.1007/s00359-023-01669-z
The seasons within: a theoretical perspective on photoperiodic entrainment and encoding
Abstract
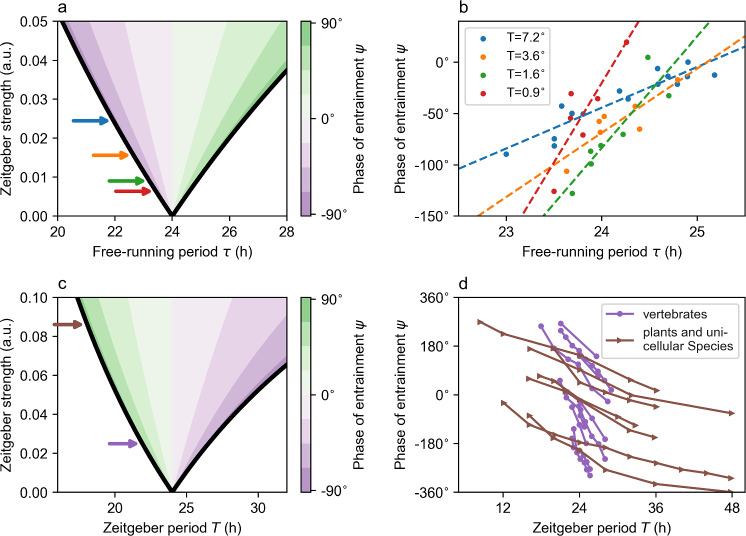
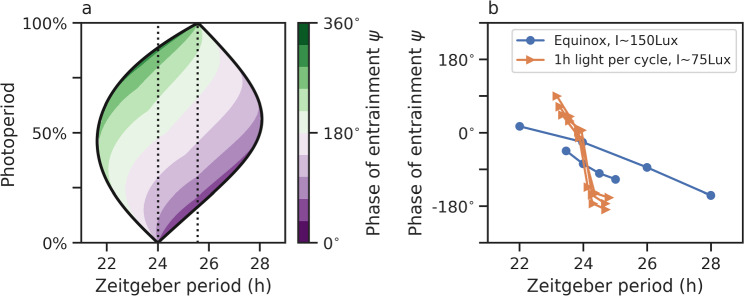
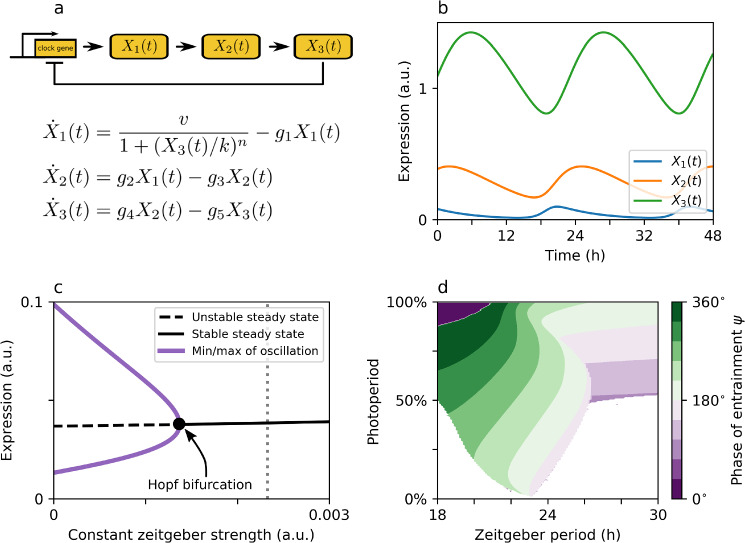
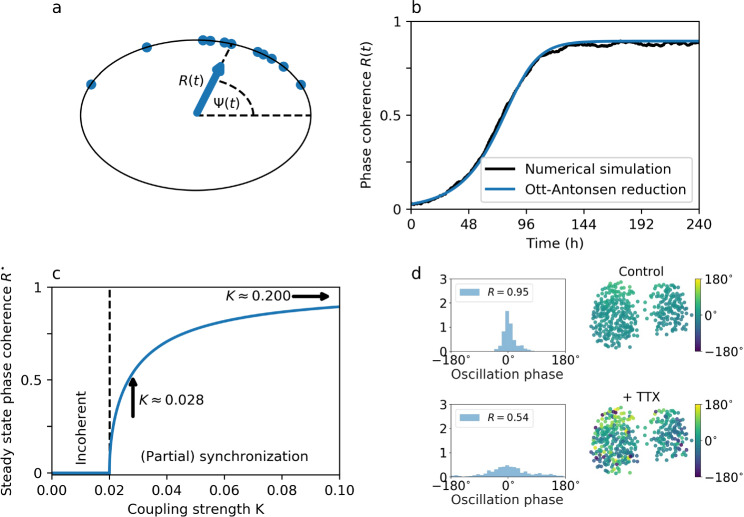
Circadian clocks are internal timing devices that have evolved as an adaption to the omnipresent natural 24 h rhythmicity of daylight intensity. Properties of the circadian system are photoperiod dependent. The phase of entrainment varies systematically with season. Plastic photoperiod-dependent re-arrangements in the mammalian circadian core pacemaker yield an internal representation of season. Output pathways of the circadian clock regulate photoperiodic responses such as flowering time in plants or hibernation in mammals. Here, we review the concepts of seasonal entrainment and photoperiodic encoding. We introduce conceptual phase oscillator models as their high level of abstraction, but, yet, intuitive interpretation of underlying parameters allows for a straightforward analysis of principles that determine entrainment characteristics. Results from this class of models are related and discussed in the context of more complex conceptual amplitude-phase oscillators as well as contextual molecular models that take into account organism, tissue, and cell-type-specific details.
Keywords: Circadian clock; Coupling; Mathematical modeling; Photoperiodic encoding; Seasonality; Synchronization.
© 2023. The Author(s).
Conflict of interest statement
The author has no competing interests to declare that are relevant to the content of this article.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
