

This is a preprint.
Insights into the Mechanisms and Structure of Breakage-Fusion-Bridge Cycles in Cervical Cancer using Long-Read Sequencing
- PMID: 37662332
- PMCID: PMC10473792
- DOI: 10.1101/2023.08.21.23294276
Insights into the Mechanisms and Structure of Breakage-Fusion-Bridge Cycles in Cervical Cancer using Long-Read Sequencing
Update in
-
Insights into the mechanisms and structure of breakage-fusion-bridge cycles in cervical cancer using long-read sequencing.Am J Hum Genet. 2024 Mar 7;111(3):544-561. doi: 10.1016/j.ajhg.2024.01.002. Epub 2024 Feb 1. Am J Hum Genet. 2024. PMID: 38307027 Free PMC article.
Abstract
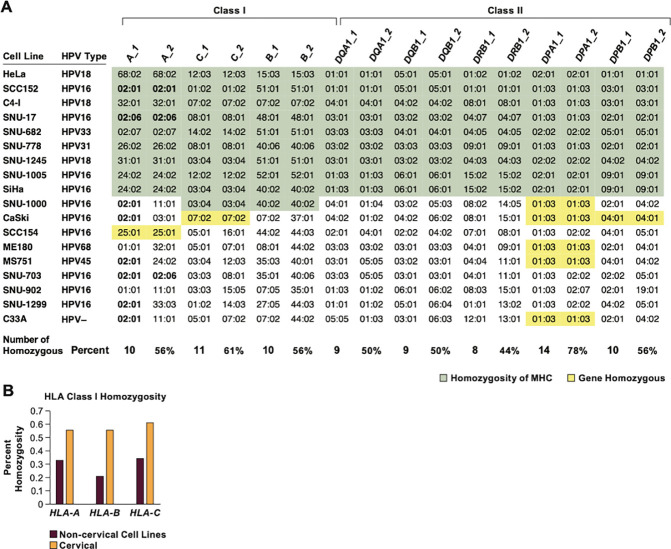
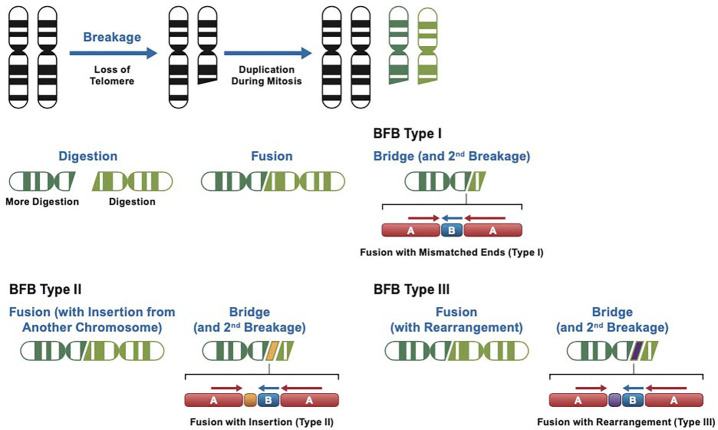
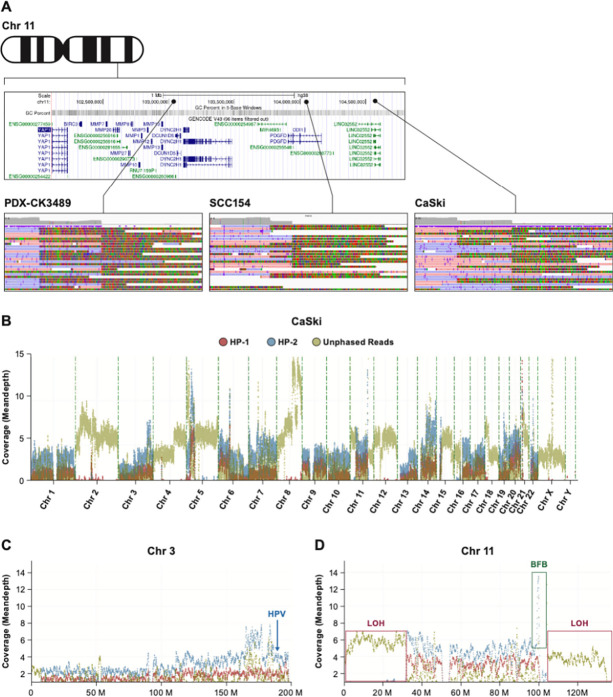
Cervical cancer is caused by human papillomavirus (HPV) infection, has few approved targeted therapeutics, and is the most common cause of cancer death in low-resource countries. We characterized 19 cervical and four head and neck cell lines using long-read DNA and RNA sequencing and identified the HPV types, HPV integration sites, chromosomal alterations, and cancer driver mutations. Structural variation analysis revealed telomeric deletions associated with DNA inversions resulting from breakage-fusion-bridge (BFB) cycles. BFB is a common mechanism of chromosomal alterations in cancer, and this is one of the first analyses of these events using long-read sequencing. Analysis of the inversion sites revealed staggered ends consistent with exonuclease digestion of the DNA after breakage. Some BFB events are complex, involving inter- or intra-chromosomal insertions or rearrangements. None of the BFB breakpoints had telomere sequences added to resolve the dicentric chromosomes and only one BFB breakpoint showed chromothripsis. Five cell lines have a Chr11q BFB event, with YAP1/BIRC2/BIRC3 gene amplification. Indeed, YAP1 amplification is associated with a 10-year earlier age of diagnosis of cervical cancer and is three times more common in African American women. This suggests that cervical cancer patients with YAP1/BIRC2/BIRC3-amplification, especially those of African American ancestry, might benefit from targeted therapy. In summary, we uncovered new insights into the mechanisms and consequences of BFB cycles in cervical cancer using long-read sequencing.
Keywords: HPV integration; Human Papillomavirus; breakage-bridge-fusion events; cervical cancer; extrachromosomal DNA.
Figures





References
-
- Andre F., Ciruelos E., Rubovszky G., Campone M., Loibl S., Rugo H. S., Iwata H., Conte P., Mayer I. A., Kaufman B., et al. (2019). Alpelisib for PIK3CA-Mutated, Hormone Receptor-Positive Advanced Breast Cancer. N Engl J Med 380, 1929–1940. - PubMed
-
- Bagutti C., Forro G., Ferralli J., Rubin B., and Chiquet-Ehrismann R. (2003). The intracellular domain of teneurin-2 has a nuclear function and represses zic-1-mediated transcription. J Cell Sci 116, 2957–2966. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous