

Hematopoietic stem and progenitor cells confer cross-protective trained immunity in mouse models
- PMID: 37664586
- PMCID: PMC10470378
- DOI: 10.1016/j.isci.2023.107596
Hematopoietic stem and progenitor cells confer cross-protective trained immunity in mouse models
Erratum in
-
Erratum: Hematopoietic stem and progenitor cells confer cross-protective trained immunity in mouse models.iScience. 2025 Feb 13;28(3):111960. doi: 10.1016/j.isci.2025.111960. eCollection 2025 Mar 21. iScience. 2025. PMID: 40201417 Free PMC article.
Abstract
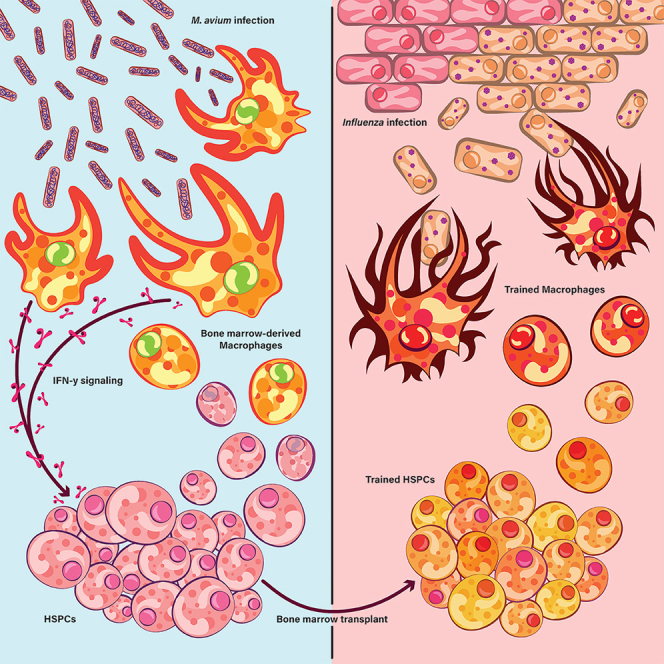
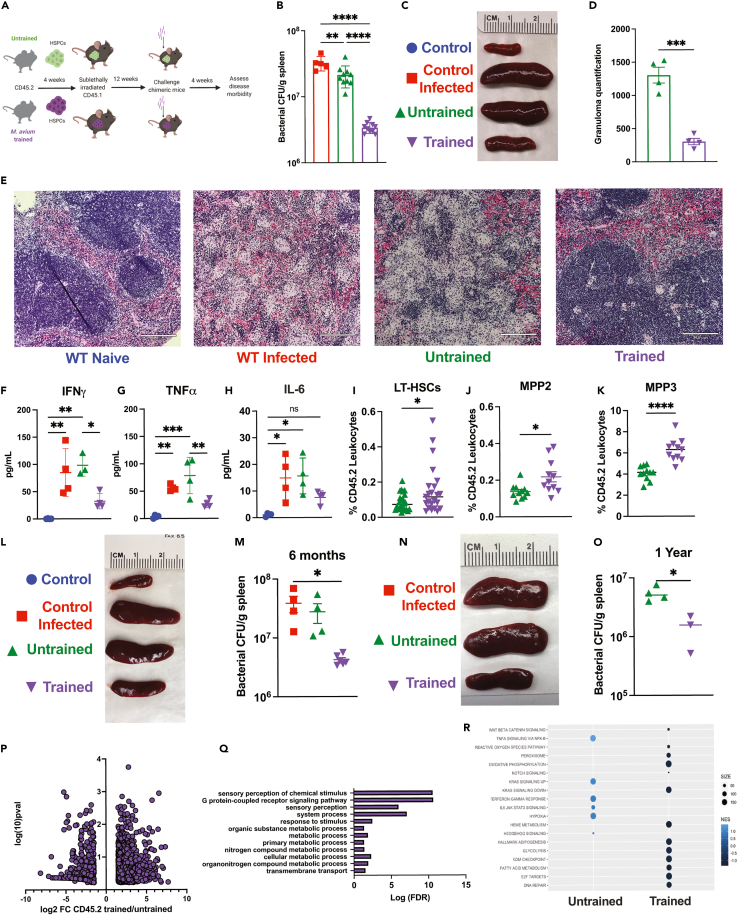
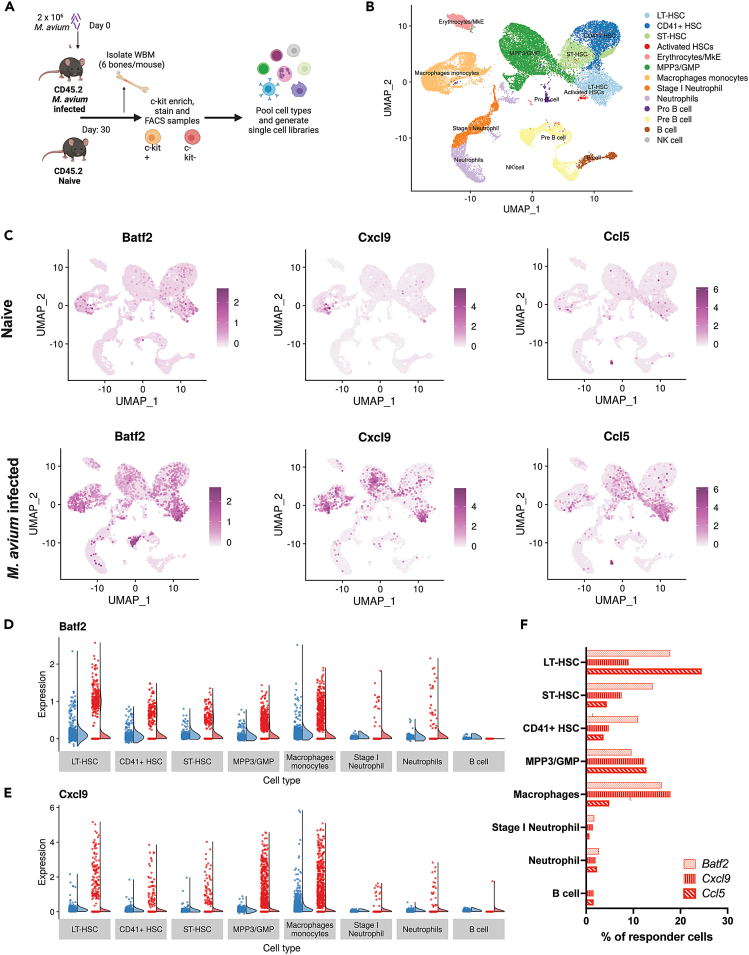
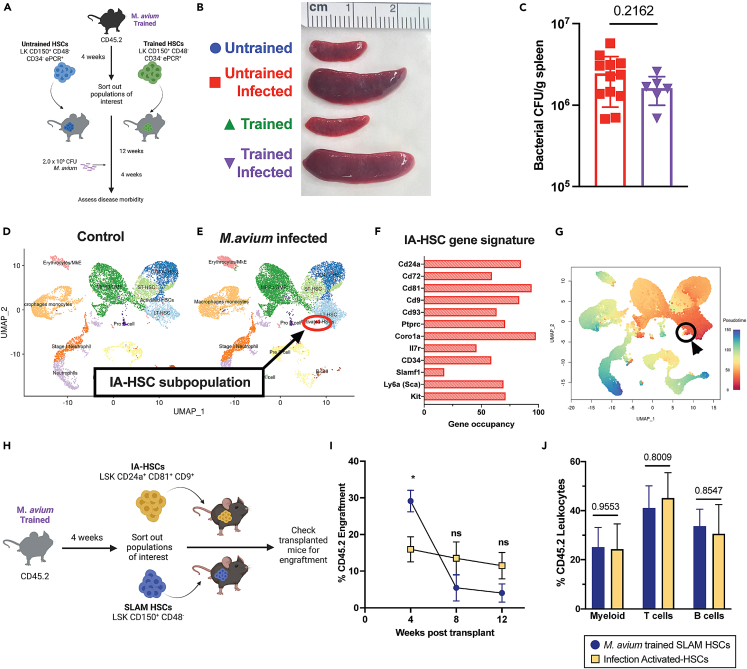
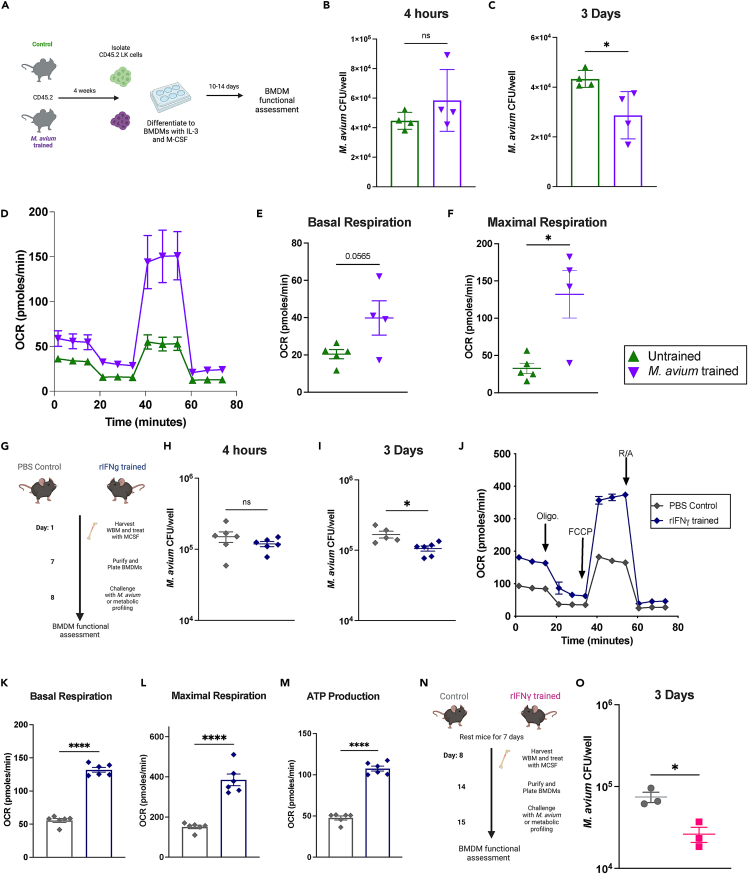
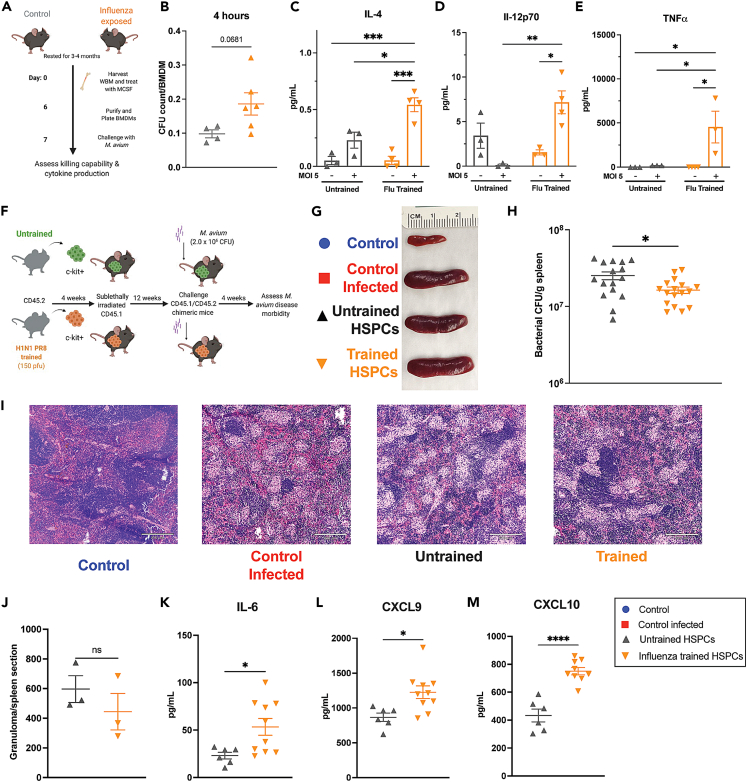
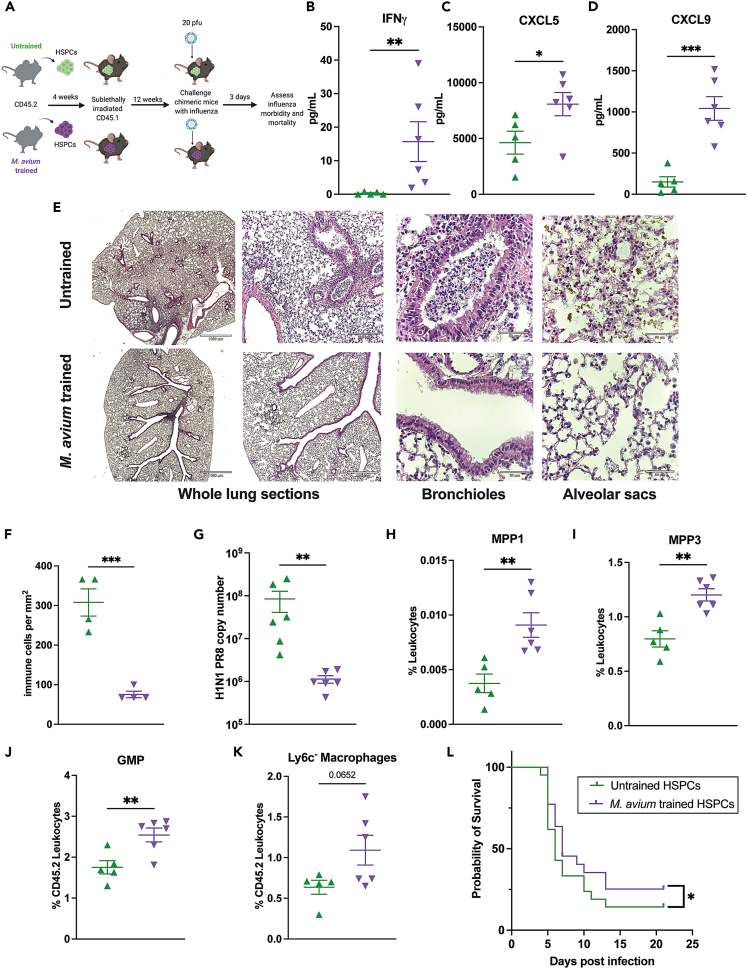
Recent studies suggest that infection reprograms hematopoietic stem and progenitor cells (HSPCs) to enhance innate immune responses upon secondary infectious challenge, a process called "trained immunity." However, the specificity and cell types responsible for this response remain poorly defined. We established a model of trained immunity in mice in response to Mycobacterium avium infection. scRNA-seq analysis revealed that HSPCs activate interferon gamma-response genes heterogeneously upon primary challenge, while rare cell populations expand. Macrophages derived from trained HSPCs demonstrated enhanced bacterial killing and metabolism, and a single dose of recombinant interferon gamma exposure was sufficient to induce similar training. Mice transplanted with influenza-trained HSPCs displayed enhanced immunity against M. avium challenge and vice versa, demonstrating cross protection against antigenically distinct pathogens. Together, these results indicate that heterogeneous responses to infection by HSPCs can lead to long-term production of bone marrow derived macrophages with enhanced function and confer cross-protection against alternative pathogens.
Keywords: Cell biology; Immunology; Microbiology; Stem cells research; Transcriptomics.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Arts R.J.W., Moorlag S.J.C.F.M., Novakovic B., Li Y., Wang S.Y., Oosting M., Kumar V., Xavier R.J., Wijmenga C., Joosten L.A.B., et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe. 2018;23:89–100.e5. doi: 10.1016/j.chom.2017.12.010. - DOI - PubMed
-
- Berendsen M.L., van Gijzel S.W., Smits J., de Mast Q., Aaby P., Benn C.S., Netea M.G., van der Ven A.J. BCG vaccination is associated with reduced malaria prevalence in children under the age of 5 years in sub-Saharan Africa. BMJ Glob. Health. 2019;4:e001862. doi: 10.1136/bmjgh-2019-001862. - DOI - PMC - PubMed
Grants and funding
- R35 HL155672/HL/NHLBI NIH HHS/United States
- S10 OD023469/OD/NIH HHS/United States
- P30 EY002520/EY/NEI NIH HHS/United States
- S10 RR024574/RR/NCRR NIH HHS/United States
- T32 DK060445/DK/NIDDK NIH HHS/United States
- R01 AI141716/AI/NIAID NIH HHS/United States
- S10 OD025240/OD/NIH HHS/United States
- R01 HL136333/HL/NHLBI NIH HHS/United States
- T32 HL092332/HL/NHLBI NIH HHS/United States
- P30 CA125123/CA/NCI NIH HHS/United States
- T32 AI053831/AI/NIAID NIH HHS/United States
- R01 HL134880/HL/NHLBI NIH HHS/United States
- R01 DK115454/DK/NIDDK NIH HHS/United States
- P01 CA265748/CA/NCI NIH HHS/United States
- F31 HL164287/HL/NHLBI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
