

Antigravitropic PIN polarization maintains non-vertical growth in lateral roots
- PMID: 37666965
- PMCID: PMC10505559
- DOI: 10.1038/s41477-023-01478-x
Antigravitropic PIN polarization maintains non-vertical growth in lateral roots
Abstract
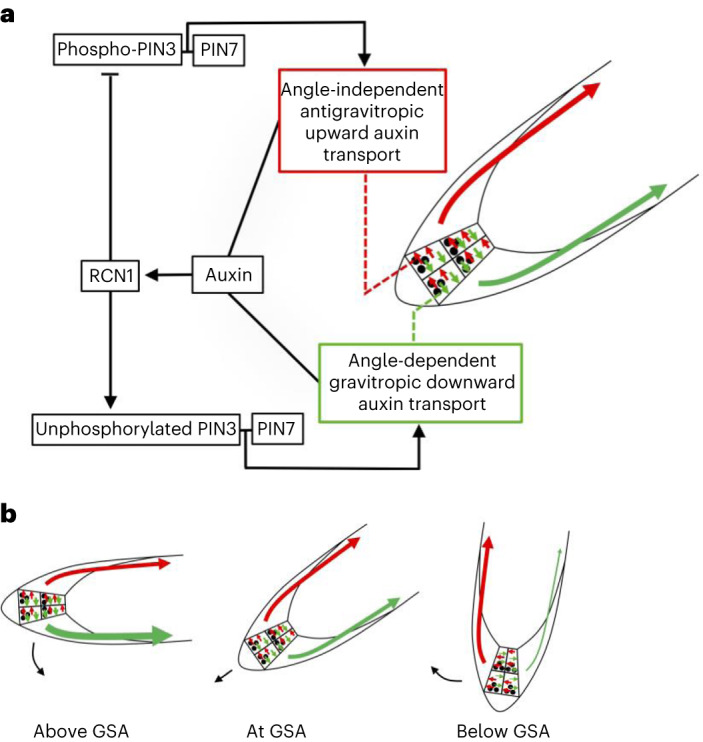
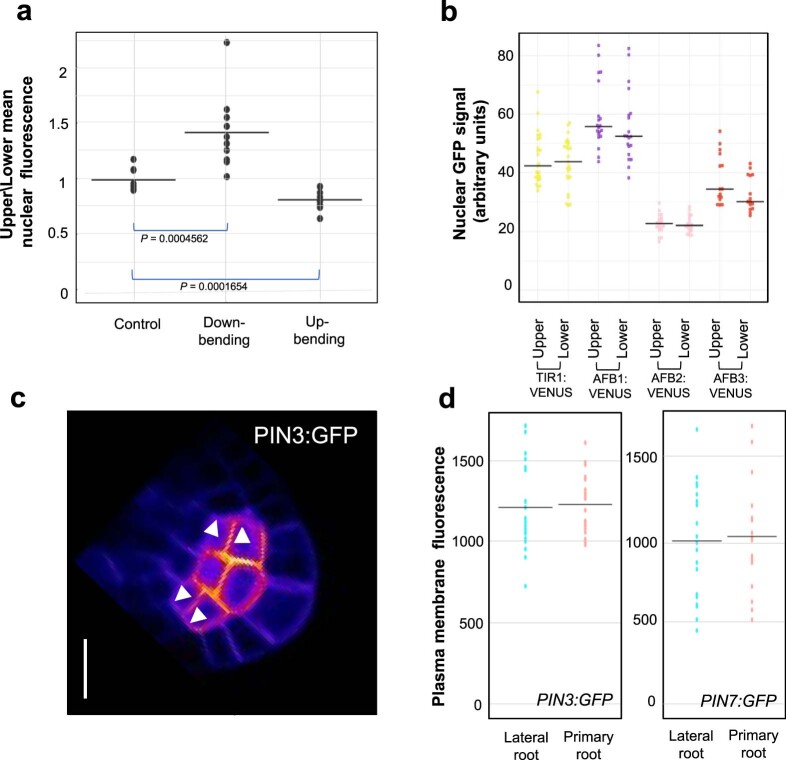
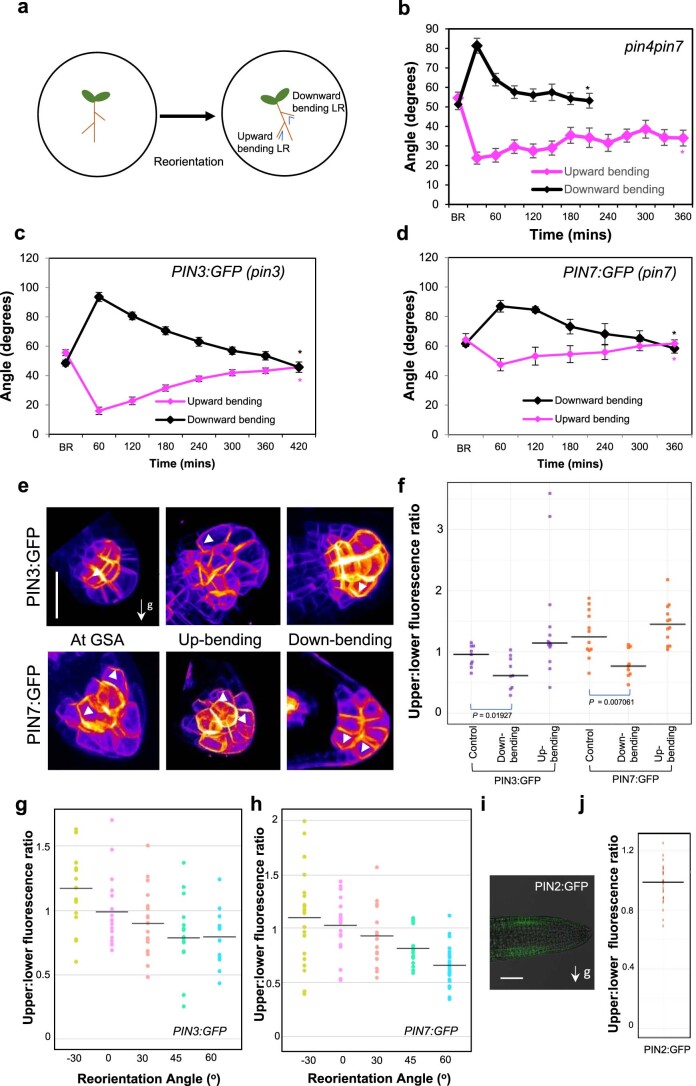
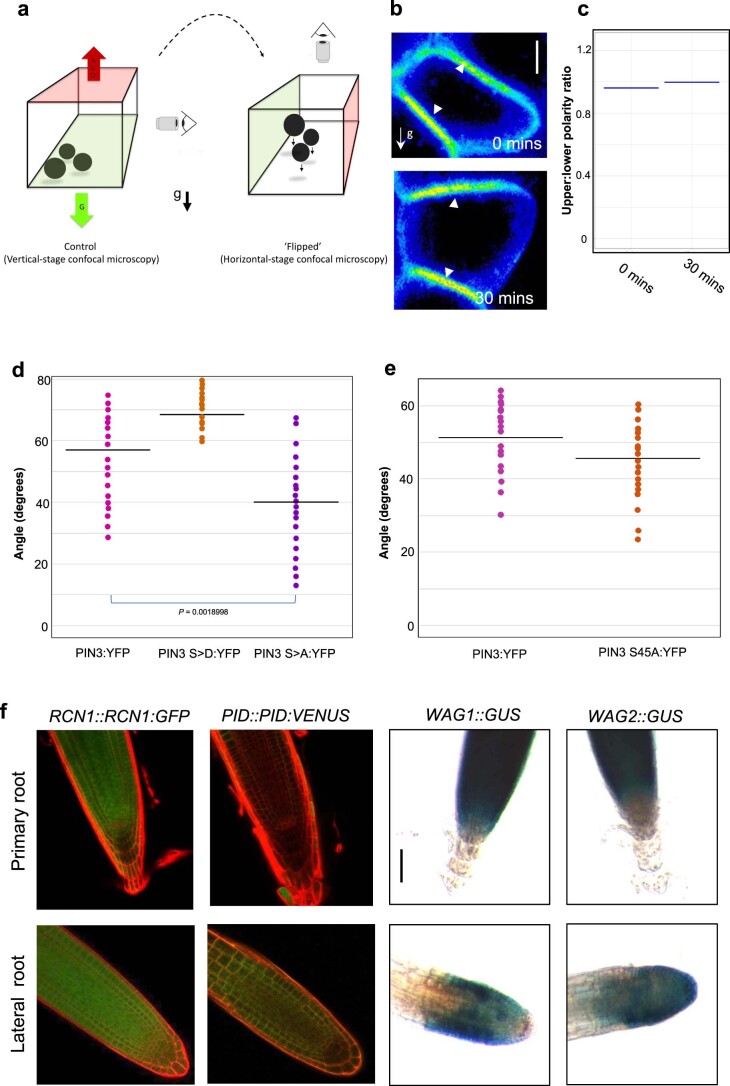
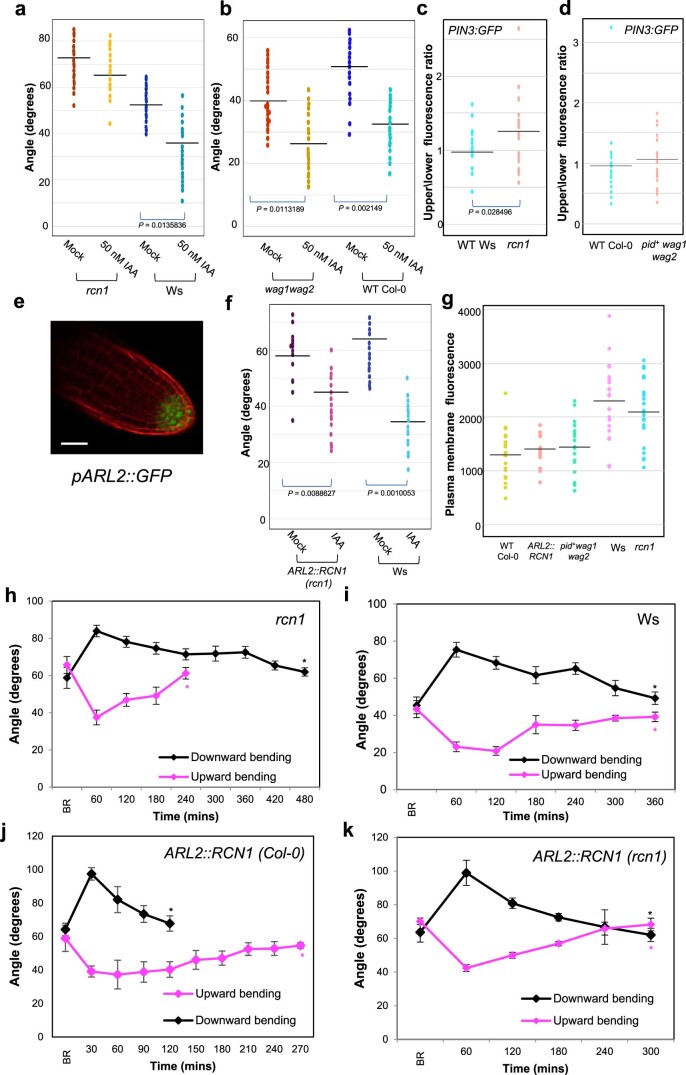
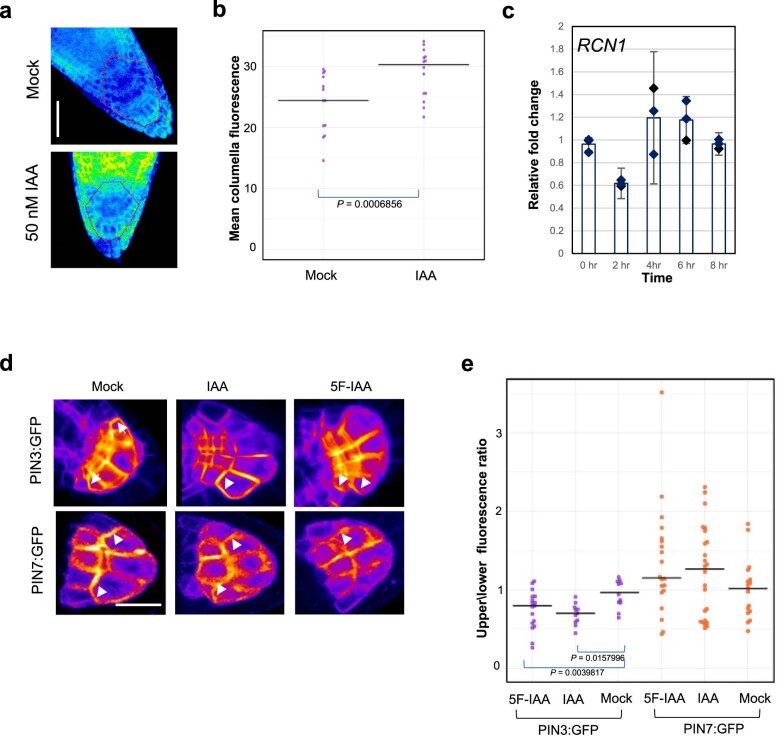
Lateral roots are typically maintained at non-vertical angles with respect to gravity. These gravitropic setpoint angles are intriguing because their maintenance requires that roots are able to effect growth response both with and against the gravity vector, a phenomenon previously attributed to gravitropism acting against an antigravitropic offset mechanism. Here we show how the components mediating gravitropism in the vertical primary root-PINs and phosphatases acting upon them-are reconfigured in their regulation such that lateral root growth at a range of angles can be maintained. We show that the ability of Arabidopsis lateral roots to bend both downward and upward requires the generation of auxin asymmetries and is driven by angle-dependent variation in downward gravitropic auxin flux acting against angle-independent upward, antigravitropic flux. Further, we demonstrate a symmetry in auxin distribution in lateral roots at gravitropic setpoint angle that can be traced back to a net, balanced polarization of PIN3 and PIN7 auxin transporters in the columella. These auxin fluxes are shifted by altering PIN protein phosphoregulation in the columella, either by introducing PIN3 phosphovariant versions or via manipulation of levels of the phosphatase subunit PP2A/RCN1. Finally, we show that auxin, in addition to driving lateral root directional growth, acts within the lateral root columella to induce more vertical growth by increasing RCN1 levels, causing a downward shift in PIN3 localization, thereby diminishing the magnitude of the upward, antigravitropic auxin flux.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Comment in
-
PIN phospho-regulation drives gravity-dependent non-vertical growth in Arabidopsis roots.Nat Plants. 2023 Sep;9(9):1383-1384. doi: 10.1038/s41477-023-01479-w. Nat Plants. 2023. PMID: 37666964 No abstract available.
References
-
- Nemec B. Ueber die Art der Wahrnehmung des Schwerkraftreizes bei den Pflanzen. Ber. Dtsch Bot. Ges. 1900;18:241–245. doi: 10.1111/j.1438-8677.1900.tb04905.x. - DOI
-
- Haberlandt G. Ueber die Perzeption des geotropischen Reizes. Ber. Dtsch Bot. Ges. 1900;18:261–272. doi: 10.1111/j.1438-8677.1900.tb04908.x. - DOI
-
- Cholodny N. Wuchshormone und Tropismem bei den Pflanzen. Biol. Zentralbl. 1927;47:604–626.
Publication types
MeSH terms
Substances
Grants and funding
- BB/N010124/1/RCUK | Biotechnology and Biological Sciences Research Council (BBSRC)
- BB/R000859/1/RCUK | Biotechnology and Biological Sciences Research Council (BBSRC)
- N/A/National Aeronautics and Space Administration (NASA)
- I 3630-B25/Austrian Science Fund (Fonds zur Förderung der Wissenschaftlichen Forschung)
- NA/Gatsby Charitable Foundation
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
