

Arrestin-3-Dependent Activation of c-Jun N-Terminal Kinases (JNKs)
- PMID: 37668419
- PMCID: PMC10624153
- DOI: 10.1002/cpz1.839
Arrestin-3-Dependent Activation of c-Jun N-Terminal Kinases (JNKs)
Abstract
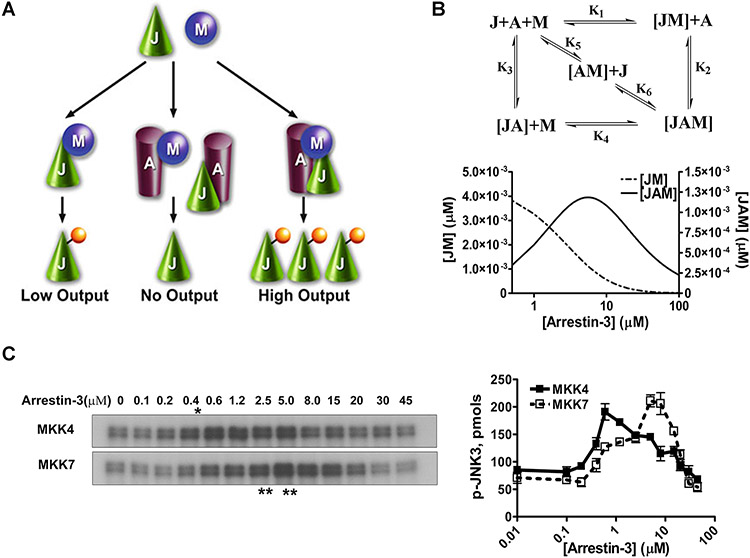
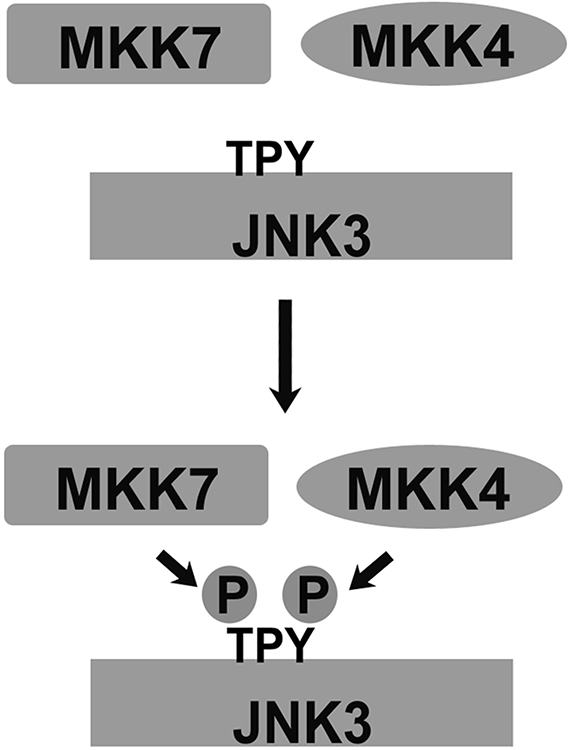
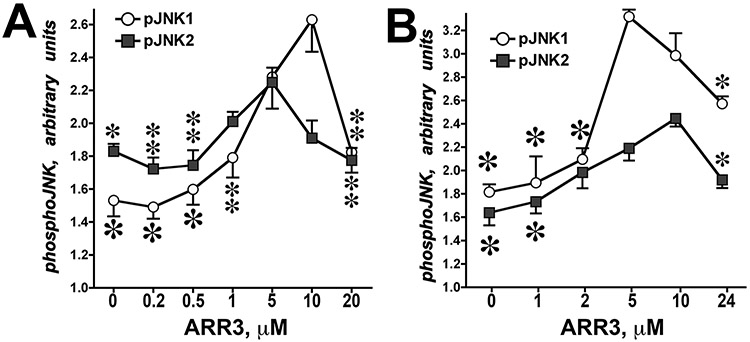
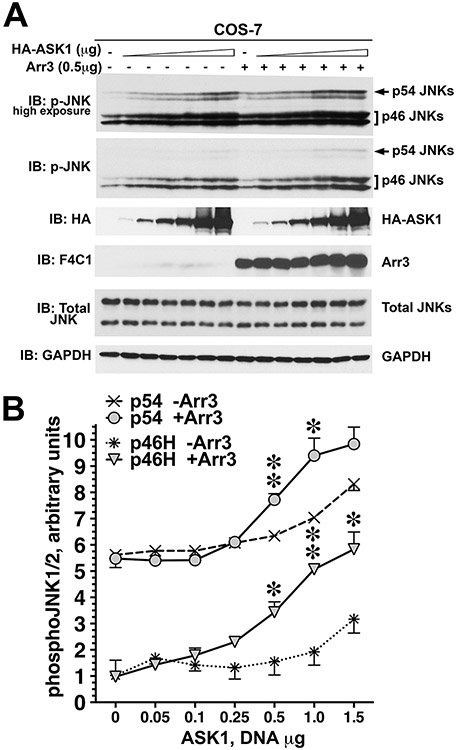
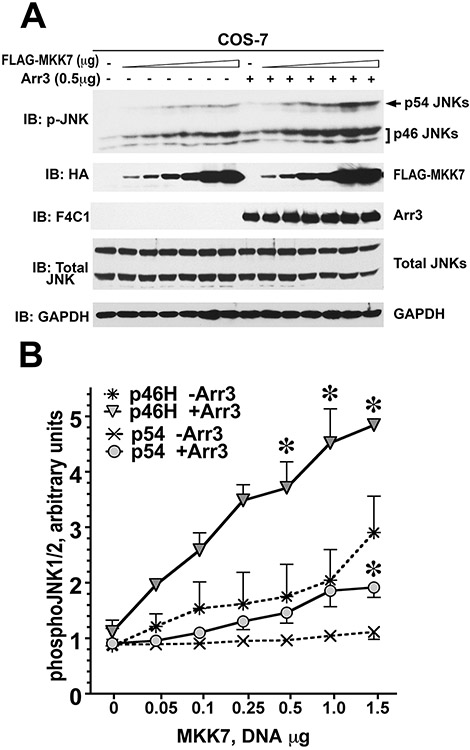
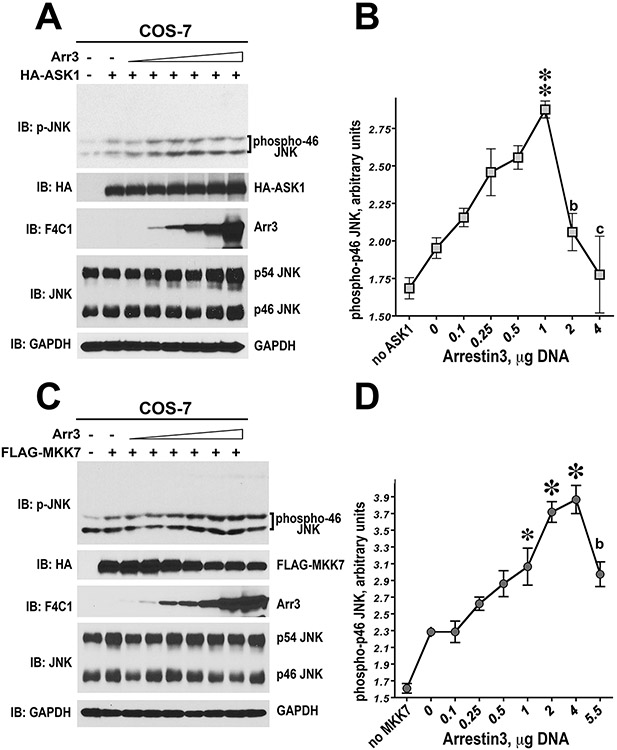
Only 1 out of 4 mammalian arrestin subtypes, arrestin-3, facilitates the activation of c-Jun N-terminal kinase (JNK) family kinases. Here, we describe two different sets of protocols used for elucidating the mechanisms involved. One is based on reconstitution of signaling modules from the following purified proteins: arrestin-3, MKK4, MKK7, JNK1, JNK2, and JNK3. The main advantage of this method is that it unambiguously establishes which effects are direct because only intended purified proteins are present in these assays. The key drawback is that the upstream-most kinases of these cascades, ASK1 or other MAP3Ks, are not available in purified form, limiting reconstitution to incomplete two-kinase modules. The other approach is used for analyzing the effects of arrestin-3 on JNK activation in intact cells. In this case, signaling modules include ASK1 and/or other MAP3Ks. However, as every cell expresses thousands of different proteins, their possible effects on the readout cannot be excluded. Nonetheless, the combination of in vitro reconstitution from purified proteins and cell-based assays makes it possible to elucidate the mechanisms of arrestin-3-dependent activation of JNK family kinases. © 2023 Wiley Periodicals LLC. Basic Protocol 1: Construction of arrestin-3-scaffolded MKK4/7-JNK1/2/3 signaling modules in vitro using purified proteins Alternate Protocol 1: Characterization of arrestin-3-mediated JNK1/2 activation by MKK4/7 by measurement of JNK1/2 phosphorylation using immunoblotting with anti-phospho-JNK antibody Support Protocol 1: Expression, purification, and activation of GST-MKK4 Support Protocol 2: Expression, purification, and activation of GST-MKK7-His6 Support Protocol 3: Expression, purification, and activation of tagless JNK1Α1 Support Protocol 4: Expression, purification, and activation of tagless JNK2Α2 Basic Protocol 2: Analysis of the role of arrestin-3 in ASK1/MKK4/MKK7-induced JNK activation in intact cells Alternate Protocol 2: Analysis of the role of arrestin-3 in MKK4-induced JNK activation in intact cells Basic Protocol 3: Characterization of the biphasic effect of arrestin-3 on ASK1/MKK7-stimulated JNK phosphorylation in cells.
Keywords: activation; arrestin; biphasic dependence; c-Jun N-terminal kinase (JNK); scaffold.
© 2023 Wiley Periodicals LLC.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
