

Ancient Clostridium DNA and variants of tetanus neurotoxins associated with human archaeological remains
- PMID: 37673908
- PMCID: PMC10482840
- DOI: 10.1038/s41467-023-41174-0
Ancient Clostridium DNA and variants of tetanus neurotoxins associated with human archaeological remains
Abstract
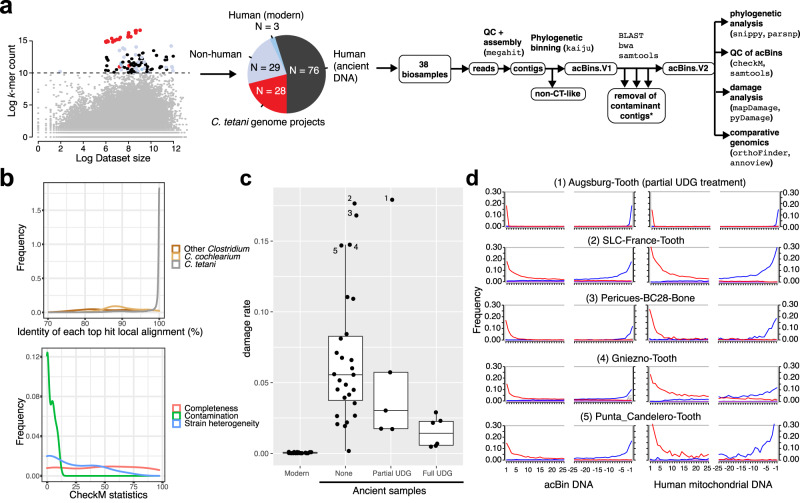
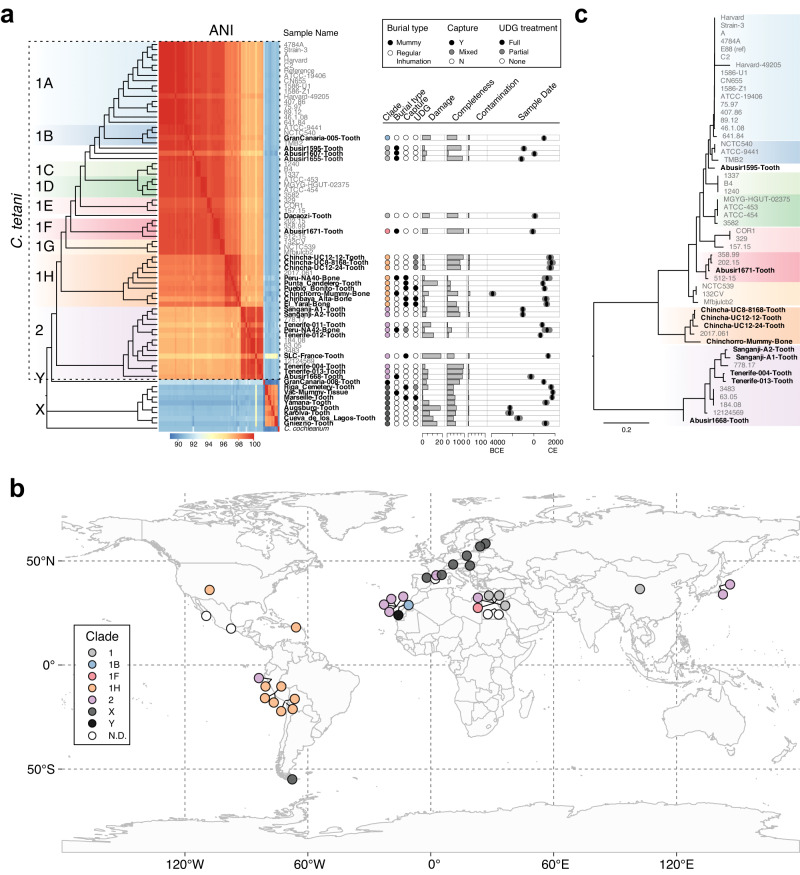
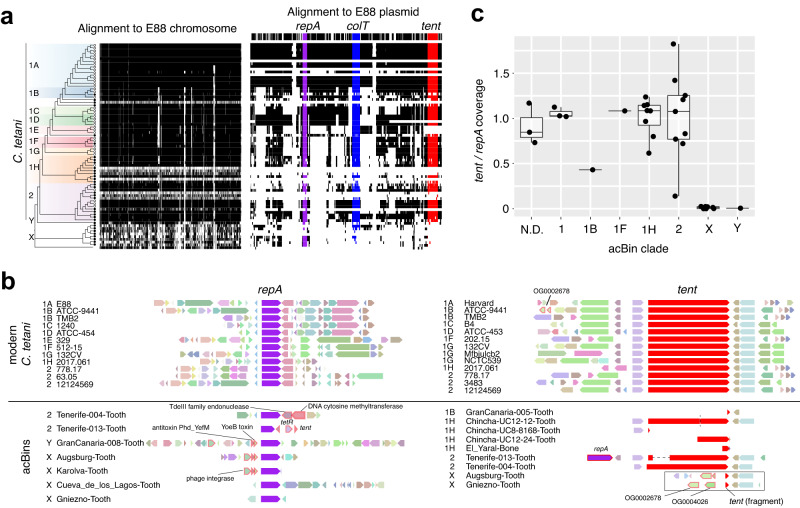
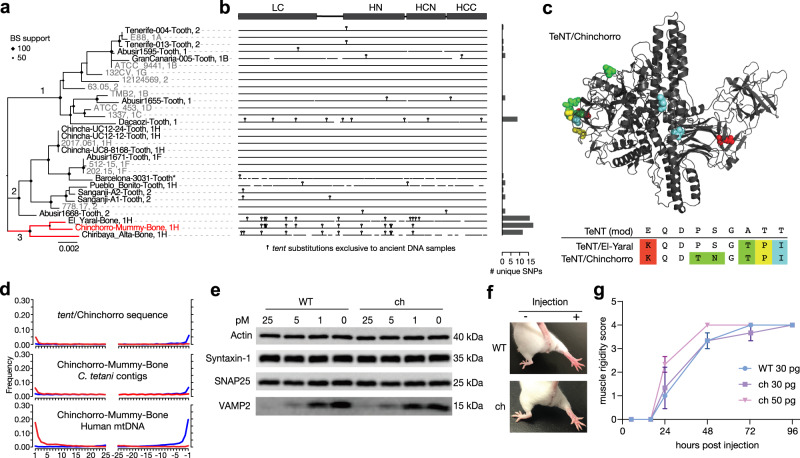
The analysis of microbial genomes from human archaeological samples offers a historic snapshot of ancient pathogens and provides insights into the origins of modern infectious diseases. Here, we analyze metagenomic datasets from 38 human archaeological samples and identify bacterial genomic sequences related to modern-day Clostridium tetani, which produces the tetanus neurotoxin (TeNT) and causes the disease tetanus. These genomic assemblies had varying levels of completeness, and a subset of them displayed hallmarks of ancient DNA damage. Phylogenetic analyses revealed known C. tetani clades as well as potentially new Clostridium lineages closely related to C. tetani. The genomic assemblies encode 13 TeNT variants with unique substitution profiles, including a subgroup of TeNT variants found exclusively in ancient samples from South America. We experimentally tested a TeNT variant selected from an ancient Chilean mummy sample and found that it induced tetanus muscle paralysis in mice, with potency comparable to modern TeNT. Thus, our ancient DNA analysis identifies DNA from neurotoxigenic C. tetani in archaeological human samples, and a novel variant of TeNT that can cause disease in mammals.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Megighian A, Pirazzini M, Fabris F, Rossetto R, Montecucco C. Tetanus and tetanus neurotoxin: From peripheral uptake to central nervous tissue targets. J. Neurochem. 2021;158:1244–1253. - PubMed
-
- Pappas G, Kiriaze IJ, Falagas ME. Insights into infectious disease in the era of Hippocrates. Int. J. Infect. Dis. 2008;12:347–350. - PubMed
-
- Miles SH. Hippocrates and informed consent. Lancet. 2009;374:1322–1323. - PubMed
-
- Sanchez, G. M. & Burridge, A. L. Decision making in head injury management in the Edwin Smith Papyrus. Neurosurg. Focus23, 1–9 (2007). - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
