

Reconciling continuous and discrete models of C4 and CAM evolution
- PMID: 37675944
- PMCID: PMC10799980
- DOI: 10.1093/aob/mcad125
Reconciling continuous and discrete models of C4 and CAM evolution
Abstract
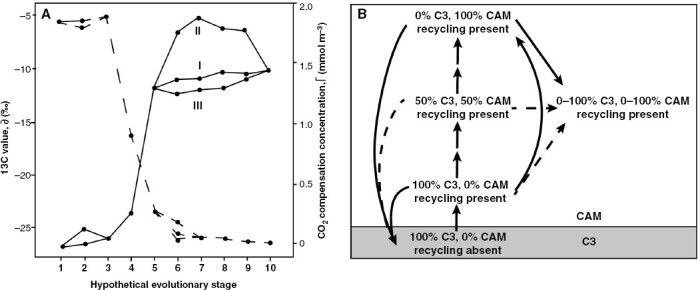
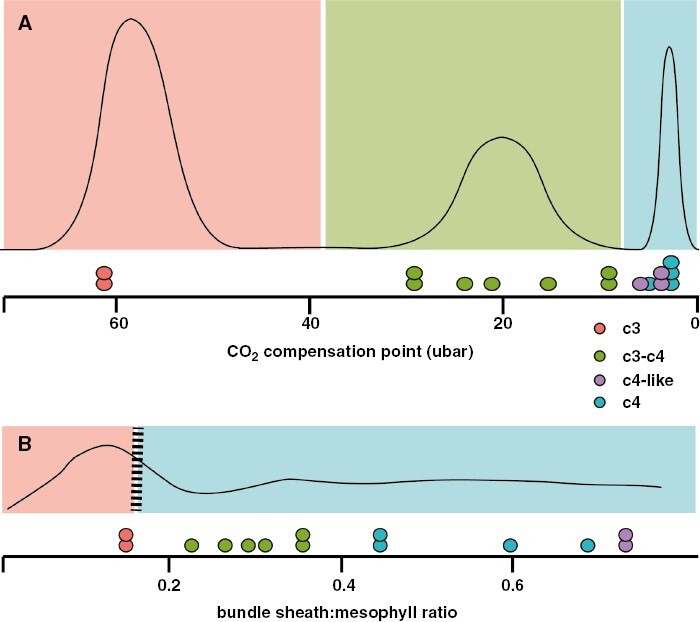
Background: A current argument in the CAM biology literature has focused on the nature of the CAM evolutionary trajectory: whether there is a smooth continuum of phenotypes between plants with C3 and CAM photosynthesis or whether there are discrete steps of phenotypic evolutionary change such as has been modelled for the evolution of C4 photosynthesis. A further implication is that a smooth continuum would increase the evolvability of CAM, whereas discrete changes would make the evolutionary transition from C3 to CAM more difficult.
Scope: In this essay, I attempt to reconcile these two viewpoints, because I think in many ways this is a false dichotomy that is constraining progress in understanding how both CAM and C4 evolved. In reality, the phenotypic space connecting C3 species and strong CAM/C4 species is both a continuum of variably expressed quantitative traits and yet also contains certain combinations of traits that we are able to identify as discrete, recognizable phenotypes. In this sense, the evolutionary mechanics of CAM origination are no different from those of C4 photosynthesis, nor from the evolution of any other complex trait assemblage.
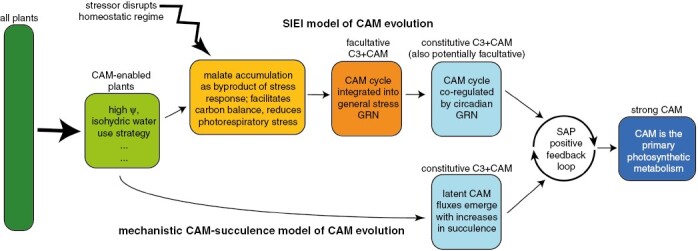
Conclusions: To make progress, we must embrace the concept of discrete phenotypic phases of CAM evolution, because their delineation will force us to articulate what aspects of phenotypic variation we think are significant. There are some current phenotypic gaps that are limiting our ability to build a complete CAM evolutionary model: the first is how a rudimentary CAM biochemical cycle becomes established, and the second is how the 'accessory' CAM cycle in C3+CAM plants is recruited into a primary metabolism. The connections to the C3 phenotype we are looking for are potentially found in the behaviour of C3 plants when undergoing physiological stress - behaviour that, strangely enough, remains essentially unexplored in this context.
Keywords: Phenotype; character evolution; continuous; discrete; heuristic.
© The Author(s) 2023. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Figures
Similar articles
-
Evolutionary trajectories, accessibility and other metaphors: the case of C4 and CAM photosynthesis.New Phytol. 2019 Sep;223(4):1742-1755. doi: 10.1111/nph.15851. Epub 2019 Jul 5. New Phytol. 2019. PMID: 30993711 Review.
-
The Road to C4 Photosynthesis: Evolution of a Complex Trait via Intermediary States.Plant Cell Physiol. 2016 May;57(5):881-9. doi: 10.1093/pcp/pcw009. Epub 2016 Feb 17. Plant Cell Physiol. 2016. PMID: 26893471 Review.
-
Modelling metabolic evolution on phenotypic fitness landscapes: a case study on C4 photosynthesis.Biochem Soc Trans. 2015 Dec;43(6):1172-6. doi: 10.1042/BST20150148. Biochem Soc Trans. 2015. PMID: 26614656 Review.
-
Are crassulacean acid metabolism and C4 photosynthesis incompatible?Funct Plant Biol. 2002 Jun;29(6):775-785. doi: 10.1071/PP01217. Funct Plant Biol. 2002. PMID: 32689525
-
Crassulacean acid metabolism: a continuous or discrete trait?New Phytol. 2015 Oct;208(1):73-8. doi: 10.1111/nph.13446. Epub 2015 May 14. New Phytol. 2015. PMID: 25975197
Cited by
-
The CAM lineages of planet Earth.Ann Bot. 2023 Nov 25;132(4):627-654. doi: 10.1093/aob/mcad135. Ann Bot. 2023. PMID: 37698538 Free PMC article.
-
One versus many independent assemblies of symbiotic nitrogen fixation in flowering plants.Nat Commun. 2025 Jun 19;16(1):5345. doi: 10.1038/s41467-025-60433-w. Nat Commun. 2025. PMID: 40537475 Free PMC article. Review.
-
CAM evolution is associated with gene family expansion in an explosive bromeliad radiation.Plant Cell. 2024 Oct 3;36(10):4109-4131. doi: 10.1093/plcell/koae130. Plant Cell. 2024. PMID: 38686825 Free PMC article.
-
Prospects and perspectives: inferring physiological and regulatory targets for CAM from molecular and modelling approaches.Ann Bot. 2023 Nov 25;132(4):583-596. doi: 10.1093/aob/mcad142. Ann Bot. 2023. PMID: 37742290 Free PMC article. Review.
-
Crassulacean acid metabolism (CAM) at the crossroads: a special issue to honour 50 years of CAM research by Klaus Winter.Ann Bot. 2023 Nov 25;132(4):553-561. doi: 10.1093/aob/mcad160. Ann Bot. 2023. PMID: 37856823 Free PMC article. No abstract available.
References
-
- Barrera Zambrano VA, Lawson T, Olmos E, Fernández-García N, Borland AM.. 2014. Leaf anatomical traits which accommodate the facultative engagement of crassulacean acid metabolism in tropical trees of the genus Clusia. Journal of Experimental Botany 65: 3513–3523. doi:10.1093/jxb/eru022. - DOI - PubMed
-
- Christin P, Sage TL, Edwards EJ, Ogburn RM, Khoshravesh R, Sage RF.. 2011. Complex evolutionary transitions and the significance of C3–C4 intermediate forms of photosynthesis in Molluginaceae. Evolution; International Journal of Organic Evolution 65: 643–660. doi:10.1111/j.1558-5646.2010.01168.x. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
