

Characterization of the SARS-CoV-2 BA.5.5 and BQ.1.1 Omicron variants in mice and hamsters
- PMID: 37676002
- PMCID: PMC10537574
- DOI: 10.1128/jvi.00628-23
Characterization of the SARS-CoV-2 BA.5.5 and BQ.1.1 Omicron variants in mice and hamsters
Abstract
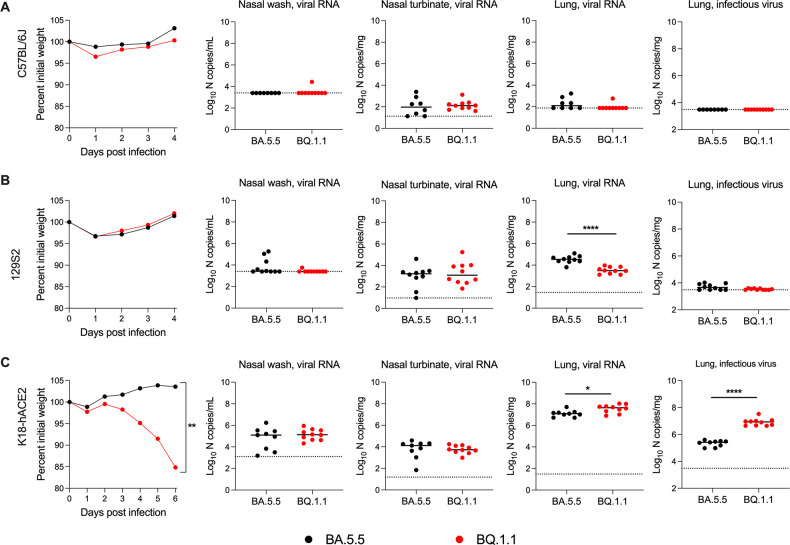
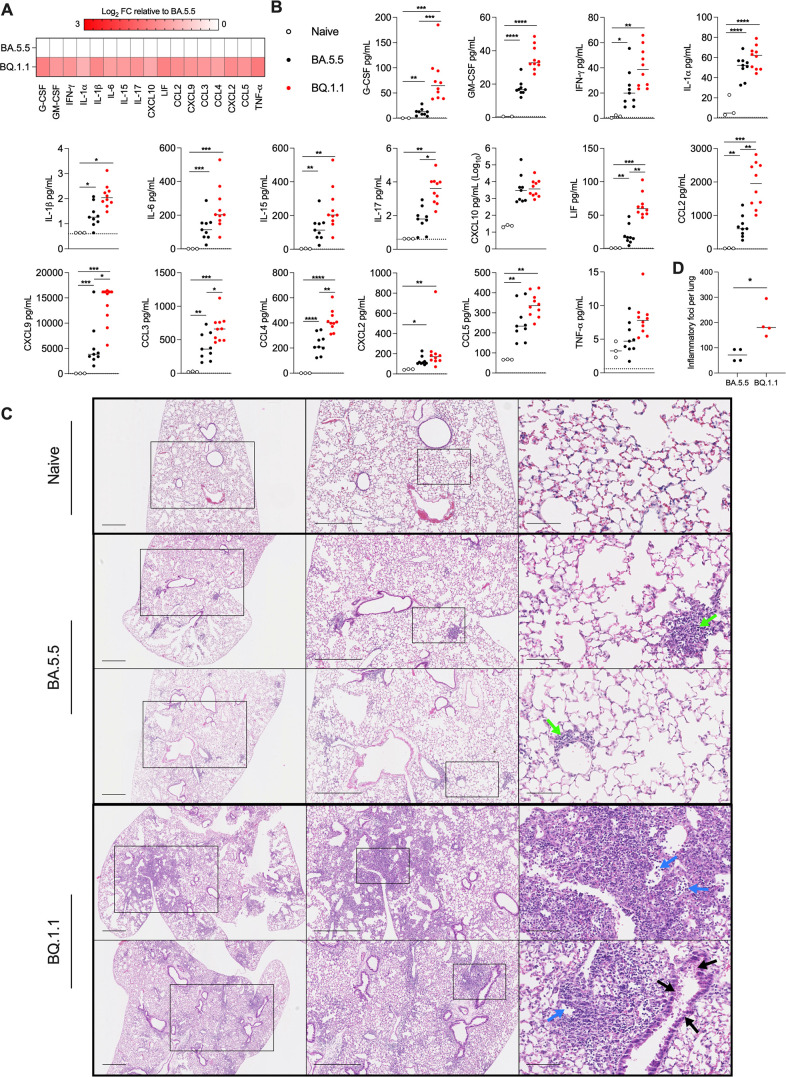
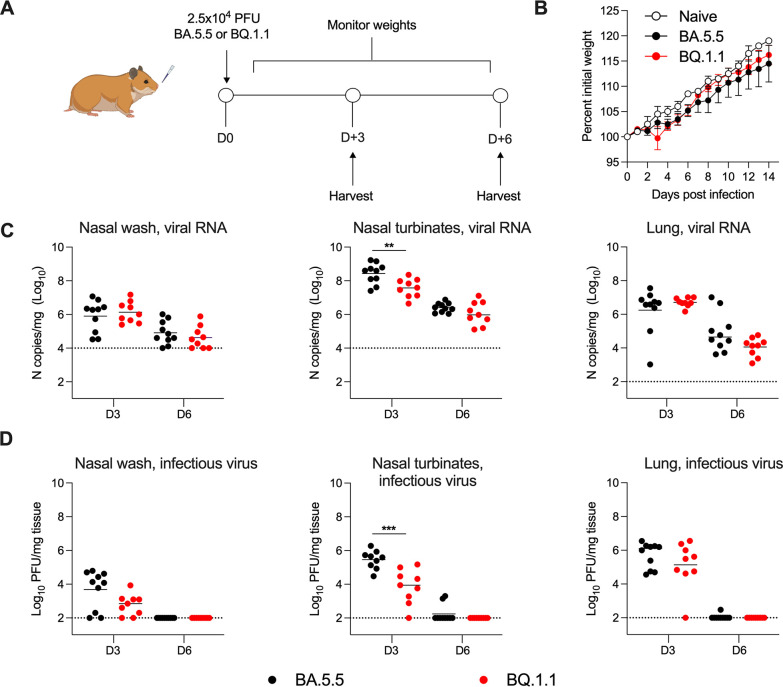
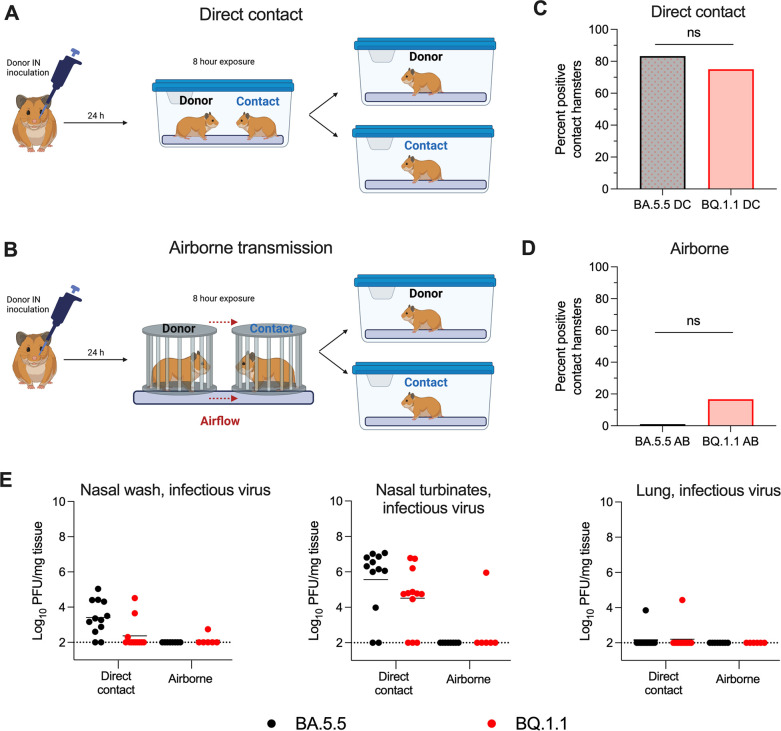
The continued evolution and emergence of novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) variants have resulted in challenges to vaccine and antibody efficacy. The emergence of each new variant necessitates the need to re-evaluate and refine animal models used for countermeasure testing. Here, we tested a recently circulating SARS-CoV-2 Omicron lineage variant, BQ.1.1, in multiple rodent models including K18-human ACE2 (hACE2) transgenic, C57BL/6J, and 129S2 mice, and Syrian golden hamsters. In contrast to a previously dominant BA.5.5 Omicron variant, inoculation of K18-hACE2 mice with BQ.1.1 resulted in substantial weight loss, a characteristic seen in pre-Omicron variants. BQ.1.1 also replicated to higher levels in the lungs of K18-hACE2 mice and caused greater lung pathology than the BA.5.5 variant. However, in C57BL/6J mice, 129S2 mice, and Syrian hamsters, BQ.1.1 did not cause increased respiratory tract infection or disease compared to animals administered BA.5.5. Moreover, the rates of direct contact or airborne transmission in hamsters were not significantly different after BQ.1.1 and BA.5.5 infections. Taken together, these data suggest that the BQ.1.1 Omicron variant has increased virulence in rodent species that express hACE2, possibly due to the acquisition of unique spike mutations relative to earlier Omicron variants. IMPORTANCE As severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) continues to evolve, there is a need to rapidly assess the efficacy of vaccines and antiviral therapeutics against newly emergent variants. To do so, the commonly used animal models must also be re-evaluated. Here, we determined the pathogenicity of the BQ.1.1 SARS-CoV-2 variant in multiple SARS-CoV-2 animal models including transgenic mice expressing human ACE2 (hACE2), two strains of conventional laboratory mice, and Syrian hamsters. While BQ.1.1 and BA.5.5 infection resulted in similar levels of viral burden and clinical disease in hamsters and the conventional strains of laboratory mice tested, increases in lung infection were detected in hACE2-expressing transgenic mice, which corresponded with greater levels of pro-inflammatory cytokines and lung pathology. Taken together, our data highlight important differences in two closely related Omicron SARS-CoV-2 variant strains and provide a foundation for evaluating countermeasures.
Keywords: BA.5.5; BQ.1.1; SARS-CoV-2; coronavirus; pathogenesis; variants.
Conflict of interest statement
M.S.D. is a consultant for Inbios, Vir Biotechnology, Ocugen, Topspin, Moderna, and Immunome. The Diamond laboratory has received unrelated funding support in sponsored research agreements from Vir Biotechnology, Emergent BioSolutions, Generate Biomedicines, and Moderna. The Boon laboratory has received unrelated funding support in sponsored research agreements from GreenLight Biosciences Inc., Moderna, and AbbVie Inc. All other authors declare no competing financial interests.
Figures
Update of
-
Characterization of the SARS-CoV-2 BA.5.5 and BQ.1.1 Omicron Variants in Mice and Hamsters.bioRxiv [Preprint]. 2023 May 1:2023.04.28.538747. doi: 10.1101/2023.04.28.538747. bioRxiv. 2023. Update in: J Virol. 2023 Sep 28;97(9):e0062823. doi: 10.1128/jvi.00628-23. PMID: 37205409 Free PMC article. Updated. Preprint.
References
-
- Halfmann PJ, Iida S, Iwatsuki-Horimoto K, Maemura T, Kiso M, Scheaffer SM, Darling TL, Joshi A, Loeber S, Singh G, Foster SL, Ying B, Case JB, Chong Z, Whitener B, Moliva J, Floyd K, Ujie M, Nakajima N, Ito M, Wright R, Uraki R, Warang P, Gagne M, Li R, Sakai-Tagawa Y, Liu Y, Larson D, Osorio JE, Hernandez-Ortiz JP, Henry AR, Ciuoderis K, Florek KR, Patel M, Odle A, Wong L-Y, Bateman AC, Wang Z, Edara V-V, Chong Z, Franks J, Jeevan T, Fabrizio T, DeBeauchamp J, Kercher L, Seiler P, Gonzalez-Reiche AS, Sordillo EM, Chang LA, van Bakel H, Simon V, Consortium Mount Sinai Pathogen Surveillance (PSP) study group, Douek DC, Sullivan NJ, Thackray LB, Ueki H, Yamayoshi S, Imai M, Perlman S, Webby RJ, Seder RA, Suthar MS, García-Sastre A, Schotsaert M, Suzuki T, Boon ACM, Diamond MS, Kawaoka Y. 2022. SARS-CoV-2 Omicron virus causes attenuated disease in mice and hamsters. Nature 603:687–692. doi: 10.1038/s41586-022-04441-6 - DOI - PMC - PubMed
-
- van Doremalen N, Avanzato VA, Goldin K, Feldmann F, Schulz JE, Haddock E, Okumura A, Lovaglio J, Hanley PW, Cordova K, Saturday G, de Wit E, Lambe T, Gilbert SC, Munster VJ. 2022. ChAdOx1 NiV vaccination protects against lethal Nipah Bangladesh virus infection in African green monkeys. NPJ Vaccines 7:171. doi: 10.1038/s41541-022-00592-9 - DOI - PMC - PubMed
-
- Hodcroft EB. 2021. Covariants: SARS-Cov-2 mutations and variants of interest. GISAID. https://covariants.org.
-
- Chen RE, Winkler ES, Case JB, Aziati ID, Bricker TL, Joshi A, Darling TL, Ying B, Errico JM, Shrihari S, VanBlargan LA, Xie X, Gilchuk P, Zost SJ, Droit L, Liu Z, Stumpf S, Wang D, Handley SA, Stine WB, Shi P-Y, Davis-Gardner ME, Suthar MS, Knight MG, Andino R, Chiu CY, Ellebedy AH, Fremont DH, Whelan SPJ, Crowe JE, Purcell L, Corti D, Boon ACM, Diamond MS. 2021. In vivo monoclonal antibody efficacy against SARS-CoV-2 variant strains. Nature 596:103–108. doi: 10.1038/s41586-021-03720-y - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
