

Electrophysiology and fluorescence to investigate cation channels and transporters in isolated plant vacuoles
- PMID: 37676514
- PMCID: PMC10442027
- DOI: 10.1007/s44154-022-00064-z
Electrophysiology and fluorescence to investigate cation channels and transporters in isolated plant vacuoles
Abstract
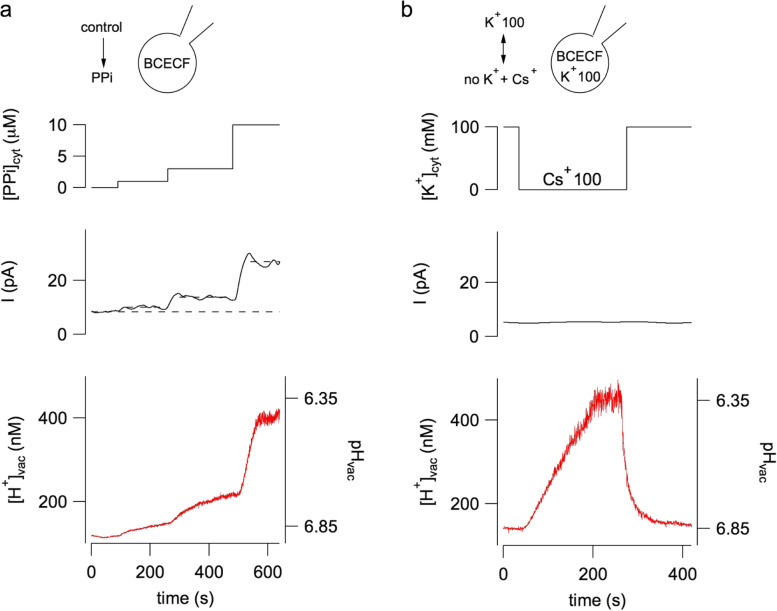
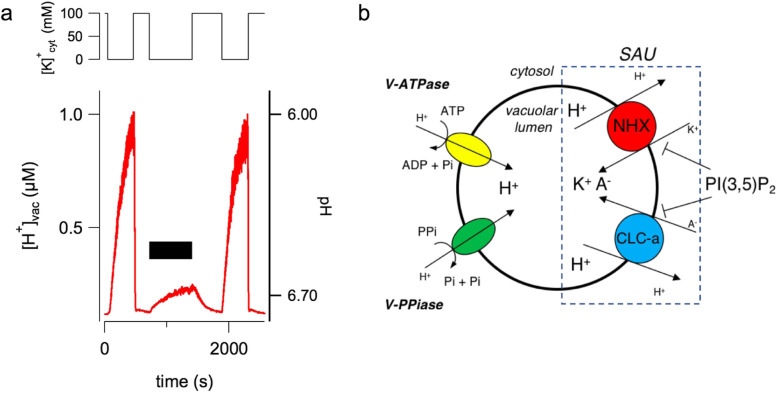
The plant vacuole plays a fundamental role in cell homeostasis. The successful application of patch-clamp technique on isolated vacuoles allows the determination of the functional characteristics of tonoplast ion channels and transporters. The parallel use of a sensor-based fluorescence approach capable of detecting changes in calcium and proton concentrations opens up new possibilities for investigation. In excised patch, the presence of fura-2 in the vacuolar solution reveals the direct permeation of calcium in plant TPC channels. In whole-vacuole, the activity of non-electrogenic NHX potassium proton antiporters can be measured by using the proton sensitive dye BCECF loaded in the vacuolar lumen by the patch pipette. Both vacuolar NHXs and CLCa (chloride/nitrate antiporter) are inhibited by the phosphoinositide PI(3,5)P2, suggesting a coordinated role of these proteins in salt accumulation. Increased knowledge in the molecular mechanisms of vacuolar ion channels and transporters has the potential to improve our understanding on how plants cope with a rapidly changing environment.
Keywords: BCECF; Calcium; Fura-2; NHX antiporters; Patch-clamp; Potassium; Proton; TPC channels.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
The phosphoinositide PI(3,5)P2 inhibits the activity of plant NHX proton/potassium antiporters: Advantages of a novel electrophysiological approach.Biomol Concepts. 2022 Mar 14;13(1):119-125. doi: 10.1515/bmc-2022-0009. Biomol Concepts. 2022. PMID: 35286784 Review.
-
Beyond the patch-clamp resolution: functional activity of nonelectrogenic vacuolar NHX proton/potassium antiporters and inhibition by phosphoinositides.New Phytol. 2021 Mar;229(5):3026-3036. doi: 10.1111/nph.17021. Epub 2020 Nov 29. New Phytol. 2021. PMID: 33098586
-
Cation Specificity of Vacuolar NHX-Type Cation/H+ Antiporters.Plant Physiol. 2019 Feb;179(2):616-629. doi: 10.1104/pp.18.01103. Epub 2018 Nov 29. Plant Physiol. 2019. PMID: 30498025 Free PMC article.
-
Fluorescence combined with excised patch: measuring calcium currents in plant cation channels.Plant J. 2009 Apr;58(1):175-82. doi: 10.1111/j.1365-313X.2008.03762.x. Epub 2009 Jan 27. Plant J. 2009. PMID: 19067975
-
How do vacuolar NHX exchangers function in plant salt tolerance?Plant Signal Behav. 2010 Jul;5(7):792-5. doi: 10.4161/psb.5.7.11767. Epub 2010 Jul 1. Plant Signal Behav. 2010. PMID: 20495345 Free PMC article. Review.
Cited by
-
Electron currents mediated by tonoplast cytochromes b561.Eur Biophys J. 2025 Aug 9. doi: 10.1007/s00249-025-01785-5. Online ahead of print. Eur Biophys J. 2025. PMID: 40782208 Review.
-
TPC1-Type Channels in Physcomitrium patens: Interaction between EF-Hands and Ca2.Plants (Basel). 2022 Dec 15;11(24):3527. doi: 10.3390/plants11243527. Plants (Basel). 2022. PMID: 36559639 Free PMC article.
References
-
- Benkerrou D, Minicozzi V, Gradogna A, Milenkovic S, Bodrenko IV, Festa M, Lagostena L, Cornara L, D’Amore A, Ceccarelli M et al (2019) A perspective on the modulation of plant and animal two pore channels (TPCs) by the flavonoid naringenin. Biophys Chem 254:106246. 10.1016/j.bpc.2019.106246 - PubMed
-
- Bertl A, Blumwald E, Coronado R, Eisenberg R, Findlay G, Gradmann D, Hille B, Köhler K, Kolb HA, MacRobbie E (1992) Electrical measurements on endomembranes. Science 258:873–874. 10.1126/science.1439795 - PubMed
-
- Boccaccio A, Scholz-Starke J, Hamamoto S, Larisch N, Festa M, Gutla PVK, Costa A, Dietrich P, Uozumi N, Carpaneto A (2014) The phosphoinositide PI (3, 5) P2 mediates activation of mammalian but not plant TPC proteins: functional expression of endolysosomal channels in yeast and plant cells. Cell Mol Life Sci 71:4275–4283. 10.1007/s00018-014-1623-2 - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
