

GWAS of random glucose in 476,326 individuals provide insights into diabetes pathophysiology, complications and treatment stratification
- PMID: 37679419
- PMCID: PMC10484788
- DOI: 10.1038/s41588-023-01462-3
GWAS of random glucose in 476,326 individuals provide insights into diabetes pathophysiology, complications and treatment stratification
Abstract
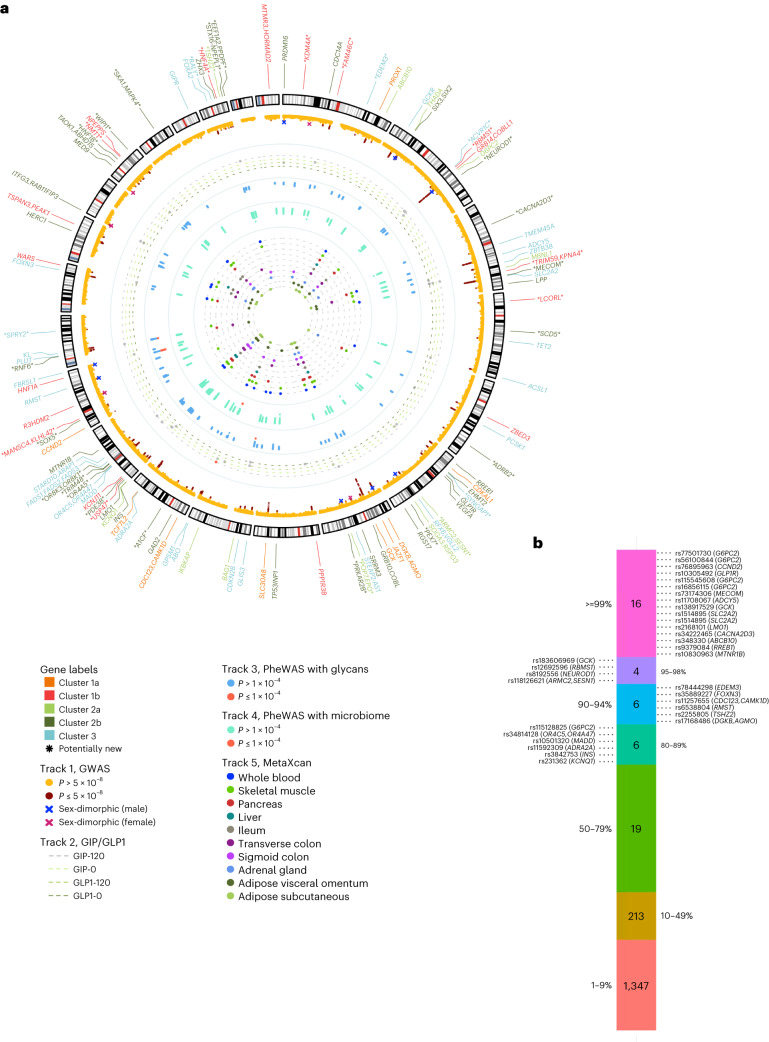
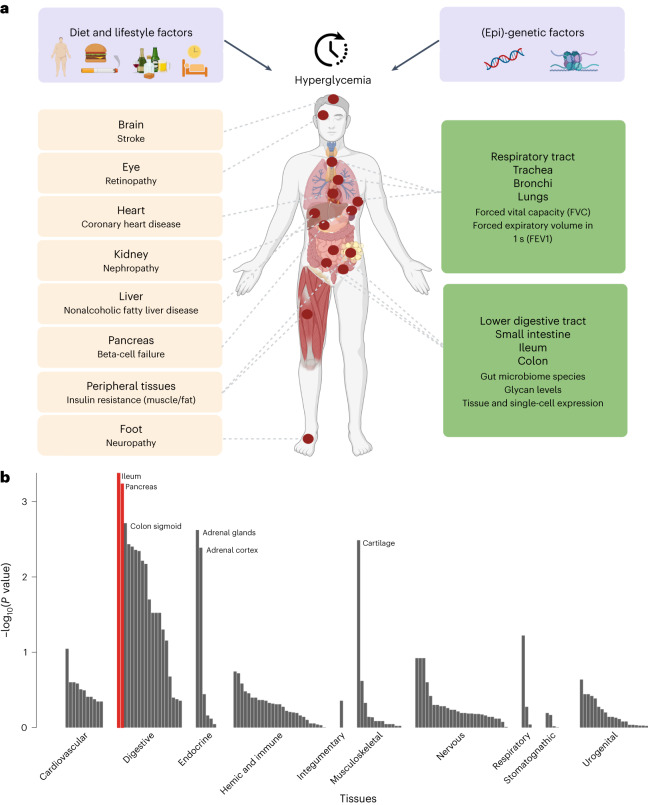
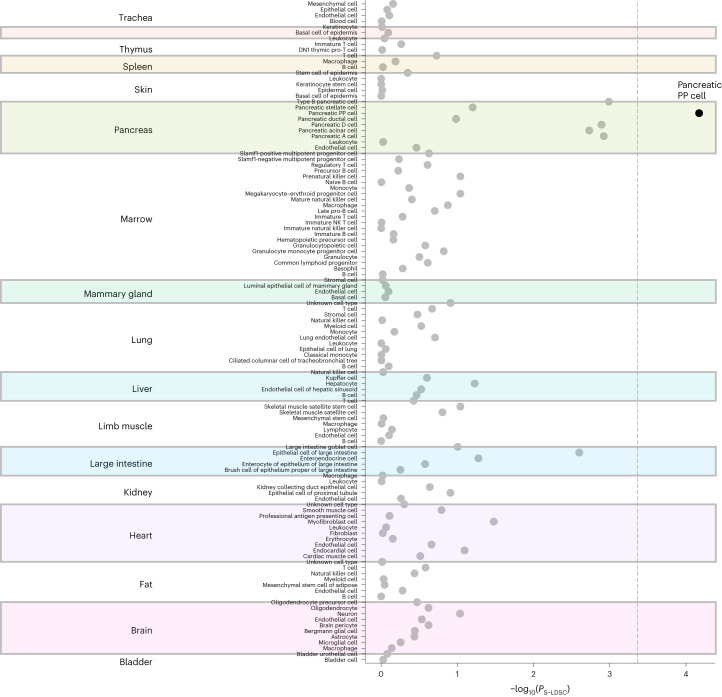
Conventional measurements of fasting and postprandial blood glucose levels investigated in genome-wide association studies (GWAS) cannot capture the effects of DNA variability on 'around the clock' glucoregulatory processes. Here we show that GWAS meta-analysis of glucose measurements under nonstandardized conditions (random glucose (RG)) in 476,326 individuals of diverse ancestries and without diabetes enables locus discovery and innovative pathophysiological observations. We discovered 120 RG loci represented by 150 distinct signals, including 13 with sex-dimorphic effects, two cross-ancestry and seven rare frequency signals. Of these, 44 loci are new for glycemic traits. Regulatory, glycosylation and metagenomic annotations highlight ileum and colon tissues, indicating an underappreciated role of the gastrointestinal tract in controlling blood glucose. Functional follow-up and molecular dynamics simulations of lower frequency coding variants in glucagon-like peptide-1 receptor (GLP1R), a type 2 diabetes treatment target, reveal that optimal selection of GLP-1R agonist therapy will benefit from tailored genetic stratification. We also provide evidence from Mendelian randomization that lung function is modulated by blood glucose and that pulmonary dysfunction is a diabetes complication. Our investigation yields new insights into the biology of glucose regulation, diabetes complications and pathways for treatment stratification.
© 2023. The Author(s).
Conflict of interest statement
A.T. has received grant funding from Sun Pharmaceuticals and Eli Lilly. J.B.M. is an academic associate for Quest Diagnostics. They make an HbA1c assay. I.R.C. is an employee of New England Biolabs, a manufacturer and vendor of reagents for life science research. M.J.C. is Chief Scientist for Genomics England, a UK Government company. The views expressed in this article are those of the author(s) and not necessarily those of the NHS, the NIHR or the Department of Health. M.I.M. has served on advisory panels for Pfizer, Novo Nordisk and Zoe Global, has received honoraria from Merck, Pfizer, Novo Nordisk and Eli Lilly and research funding from Abbvie, AstraZeneca, Boehringer Ingelheim, Eli Lilly, Janssen, Merck, Novo Nordisk, Pfizer, Roche, Sanofi Aventis, Servier and Takeda. As of June 2019, M.I.M. is an employee of Genentech and a holder of Roche stock. P.M.S. received grant funding from Laboratoires Servier. P.M.S. and D.W. receive funding from Astex Pharmaceuticals and Novo Nordisk. They are both shareholders of Septerna, where P.M.S. is also a founder. P.M.S. is the director and D.W. the Monash Node leader of the Australian Research Council of Australia Center for Cryo-Electron Microscopy of Membrane Proteins that includes the following as Partner Organizations who provide cash or in-kind funding: Astex Pharmaceuticals, AstraZeneca, Boehringer Ingelheim, Catalyst Therapeutics, Dimerix Bioscience, Genentech, Novo Nordisk, Pfizer, Sanofi Aventis, Servier and Thermo Fisher Scientific. T.J. is now a GSK employee. W. Koenig reports consulting fees from AstraZeneca, Novartis, Pfizer, The Medicines Company, DalCor, Kowa, Amgen, Corvidia, Daiichi-Sankyo, Genentech, Novo Nordisk, Esperion, OMEICOS, LIB Therapeutics; speaker honoraria from Amgen, Novartis, Berlin-Chemie, Sanofi and Bristol-Myers Squibb; grants and nonfinancial support from Abbott, Roche Diagnostics, Beckmann and Singulex, all outside the submitted work. Y.S.A. is the owner of Maatschap PolyOmica and PolyKnomics BV, private organizations providing services, research and development in the field of computational and statistical, quantitative and computational (gen)omics. G.T., U.T. and K.S. are employees of deCODE genetics/Amgen. The other authors declare no competing interests.
Figures









References
Publication types
MeSH terms
Substances
Grants and funding
- CH/1992001/6764/BHF_/British Heart Foundation/United Kingdom
- MR/N003284/1/MRC_/Medical Research Council/United Kingdom
- R01 DK072193/DK/NIDDK NIH HHS/United States
- MC_PC_14089/MRC_/Medical Research Council/United Kingdom
- MR/S003061/1/MRC_/Medical Research Council/United Kingdom
- MC_EX_MR/M009203/1/MRC_/Medical Research Council/United Kingdom
- MR/R023484/1/MRC_/Medical Research Council/United Kingdom
- MC_UU_00006/1/MRC_/Medical Research Council/United Kingdom
- MR/R010676/1/MRC_/Medical Research Council/United Kingdom
- SP/19/2/34462/BHF_/British Heart Foundation/United Kingdom
- MR/M009203/1/MRC_/Medical Research Council/United Kingdom
- MR/L01341X/1/MRC_/Medical Research Council/United Kingdom
- R01 DK062370/DK/NIDDK NIH HHS/United States
- U01 DK105535/DK/NIDDK NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
LinkOut - more resources
Full Text Sources
Medical
