

The evolution and heterogeneity of neutrophils in cancers: origins, subsets, functions, orchestrations and clinical applications
- PMID: 37679744
- PMCID: PMC10483725
- DOI: 10.1186/s12943-023-01843-6
The evolution and heterogeneity of neutrophils in cancers: origins, subsets, functions, orchestrations and clinical applications
Abstract
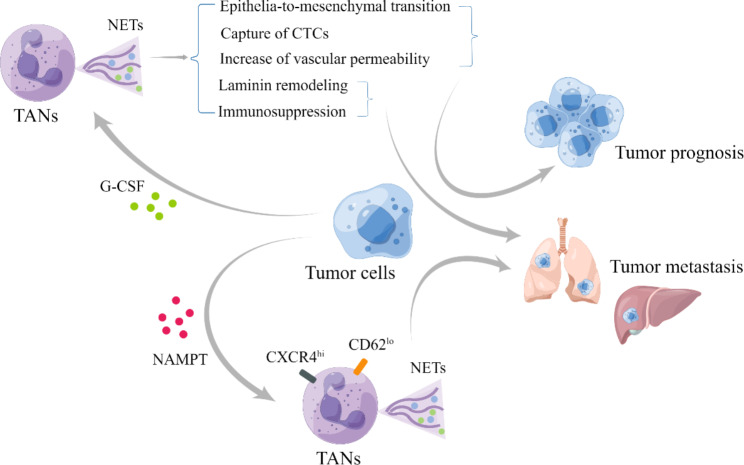
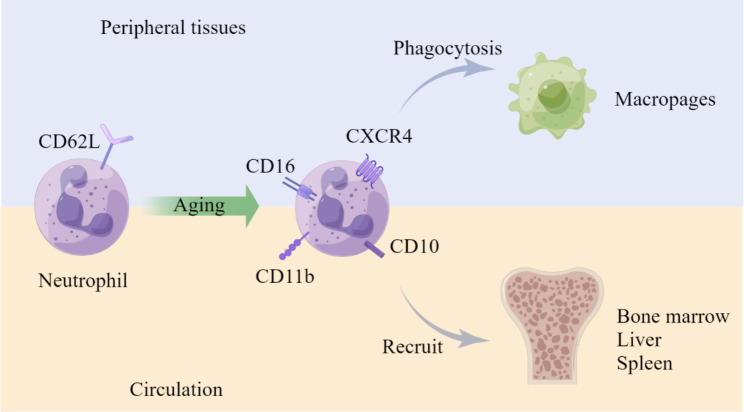
Neutrophils, the most prevalent innate immune cells in humans, have garnered significant attention in recent years due to their involvement in cancer progression. This comprehensive review aimed to elucidate the important roles and underlying mechanisms of neutrophils in cancer from the perspective of their whole life cycle, tracking them from development in the bone marrow to circulation and finally to the tumor microenvironment (TME). Based on an understanding of their heterogeneity, we described the relationship between abnormal neutrophils and clinical manifestations in cancer. Specifically, we explored the function, origin, and polarization of neutrophils within the TME. Furthermore, we also undertook an extensive analysis of the intricate relationship between neutrophils and clinical management, including neutrophil-based clinical treatment strategies. In conclusion, we firmly assert that directing future research endeavors towards comprehending the remarkable heterogeneity exhibited by neutrophils is of paramount importance.
Keywords: Clinical applications; Functions; Heterogeneity; Neutrophils; Orchestrations; Origins; Subsets; TANs (tumor-associated neutrophils).
© 2023. BioMed Central Ltd., part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
The authors have no conflict of interest.
Figures
References
-
- Hidalgo A, Chilvers ER, Summers C, Koenderman L. The Neutrophil Life Cycle. Trends Immunol. 2019;40:584–97. - PubMed
-
- Kolaczkowska E, Kubes P. Neutrophil recruitment and function in health and inflammation. Nat Rev Immunol. 2013;13:159–75. - PubMed
-
- Pillay J, den Braber I, Vrisekoop N, Kwast LM, de Boer RJ, Borghans JA, Tesselaar K, Koenderman L. In vivo labeling with 2H2O reveals a human neutrophil lifespan of 5.4 days. Blood. 2010;116:625–7. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
