

Small intestine vs. colon ecology and physiology: Why it matters in probiotic administration
- PMID: 37683651
- PMCID: PMC10518632
- DOI: 10.1016/j.xcrm.2023.101190
Small intestine vs. colon ecology and physiology: Why it matters in probiotic administration
Abstract
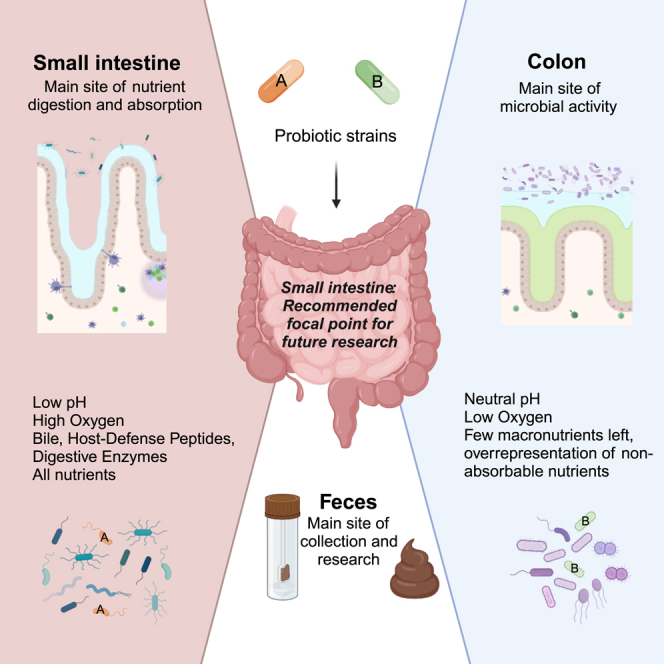
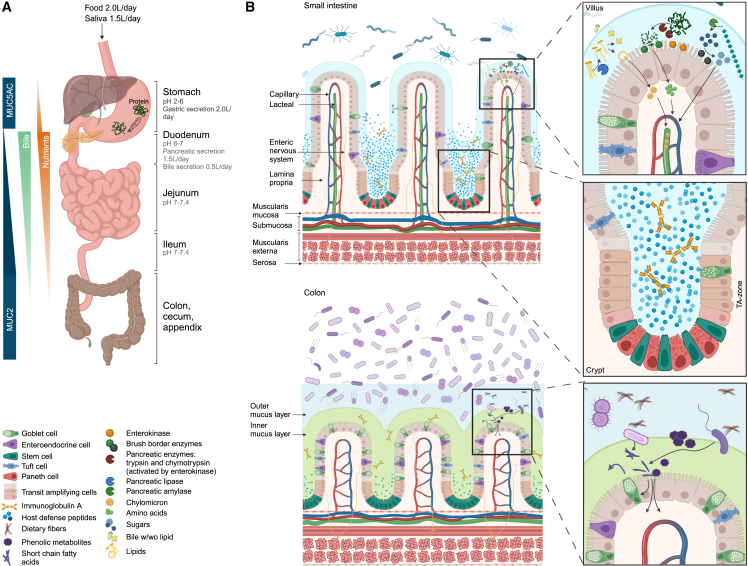
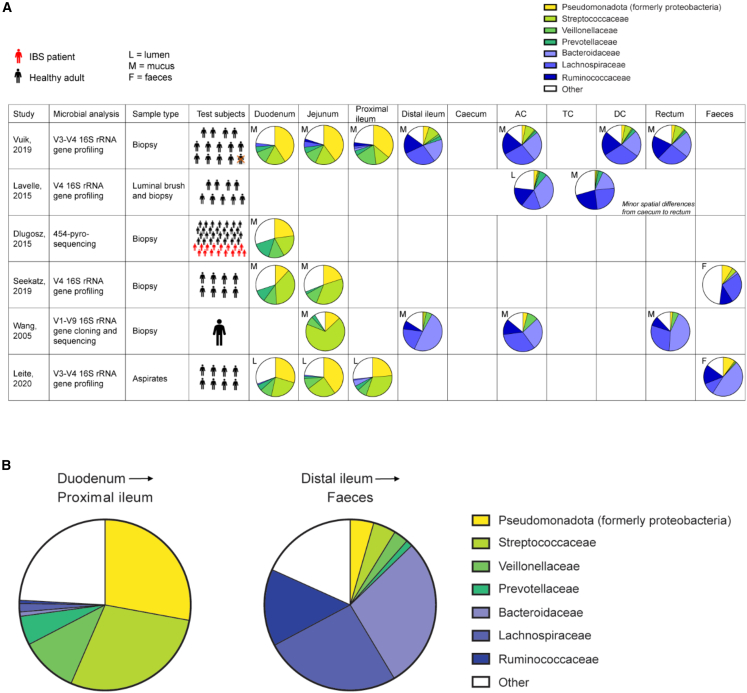
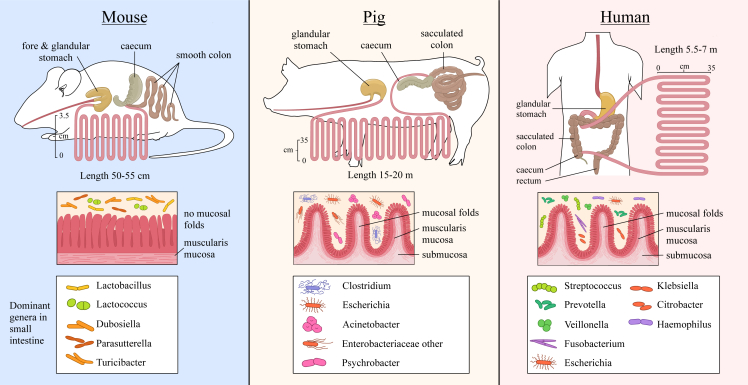
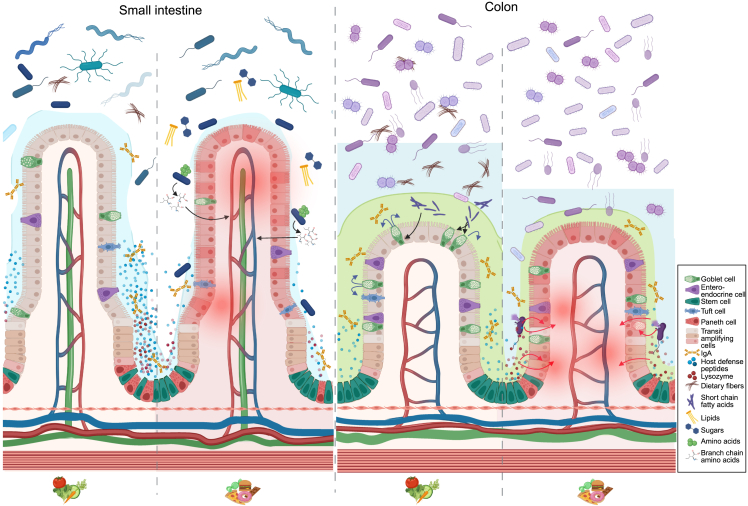
Research on gut microbiota has generally focused on fecal samples, representing luminal content of the large intestine. However, nutrient uptake is restricted to the small intestine. Abundant immune cell populations at this anatomical site combined with diminished mucus secretion and looser junctions (partly to allow for more efficient fluid and nutrient absorption) also results in intimate host-microbe interactions despite more rapid transit. It is thus crucial to dissect key differences in both ecology and physiology between small and large intestine to better leverage the immense potential of human gut microbiota imprinting, including probiotic engraftment at biological sensible niches. Here, we provide a detailed review unfolding how the physiological and anatomical differences between the small and large intestine affect gut microbiota composition, function, and plasticity. This information is key to understanding how gut microbiota manipulation, including probiotic administration, may strain-dependently transform host-microbe interactions at defined locations.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.C.O., M.N., B.P., D.S., L.G.W.S., and P.V.D.A. work full time for IFF, ADM, Yakult, Novozymes, Caelus Health, and Cryptobiotix respectively. These companies were not involved in carrying out this research.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
