

CircXRN2 suppresses tumor progression driven by histone lactylation through activating the Hippo pathway in human bladder cancer
- PMID: 37684641
- PMCID: PMC10486081
- DOI: 10.1186/s12943-023-01856-1
CircXRN2 suppresses tumor progression driven by histone lactylation through activating the Hippo pathway in human bladder cancer
Abstract
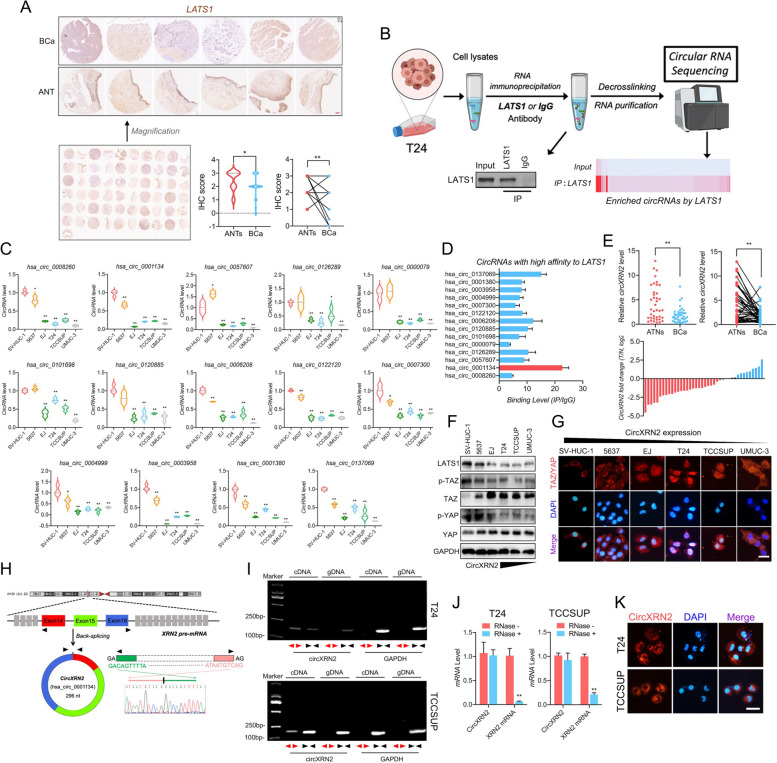
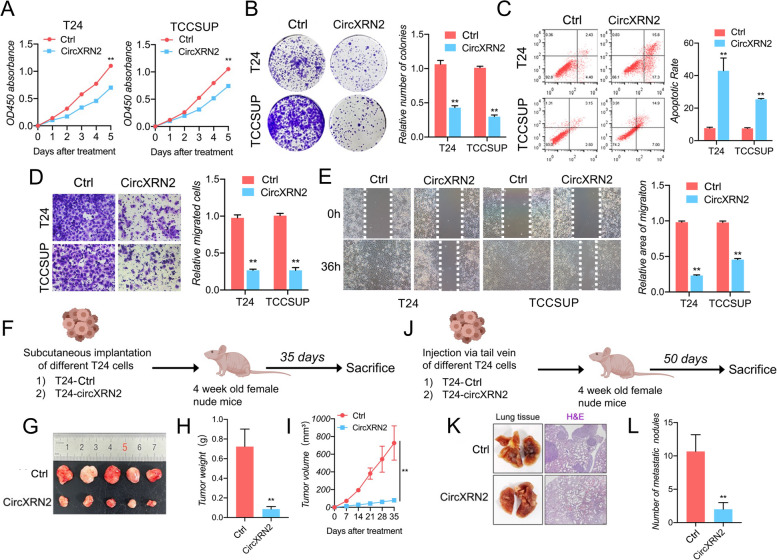
Background: Bladder cancer (BCa) is the fourth most common malignant tumor with a poor prognosis worldwide. Further exploration and research are needed to unmask the underlying roles and molecular mechanisms of circular RNAs. In the current study, our findings showed that circXRN2 suppresses tumor progression driven by histone lactylation by activating the Hippo pathway in human bladder cancer.
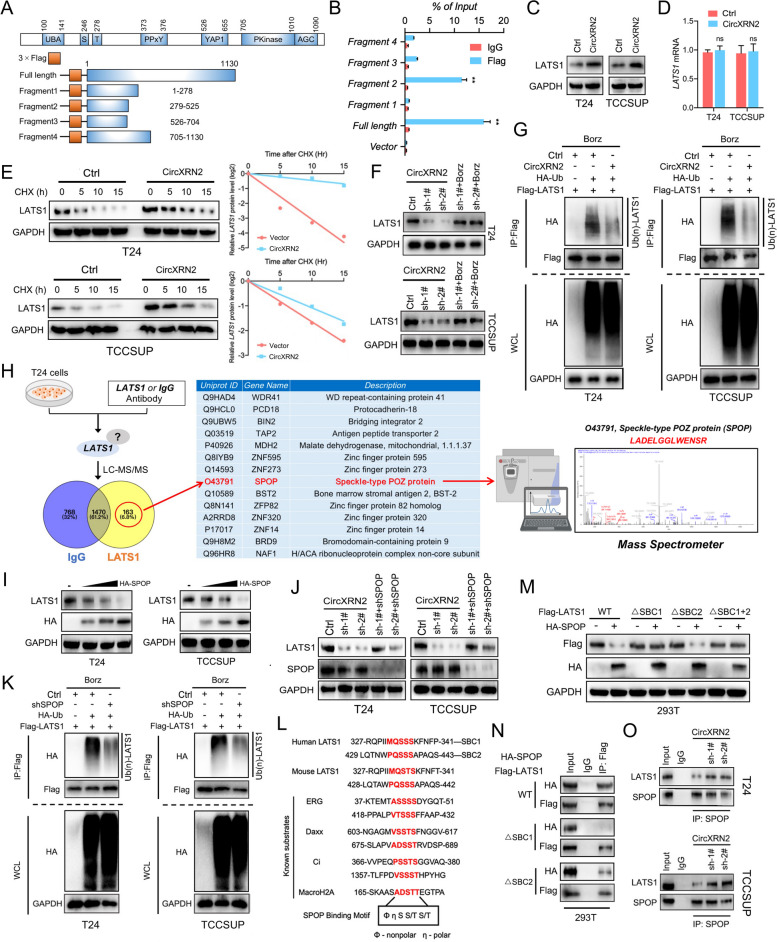
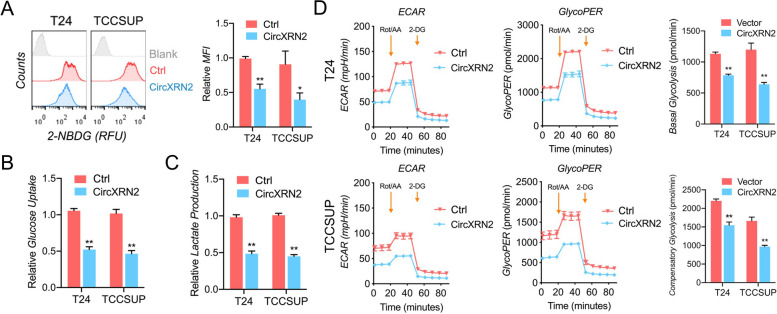
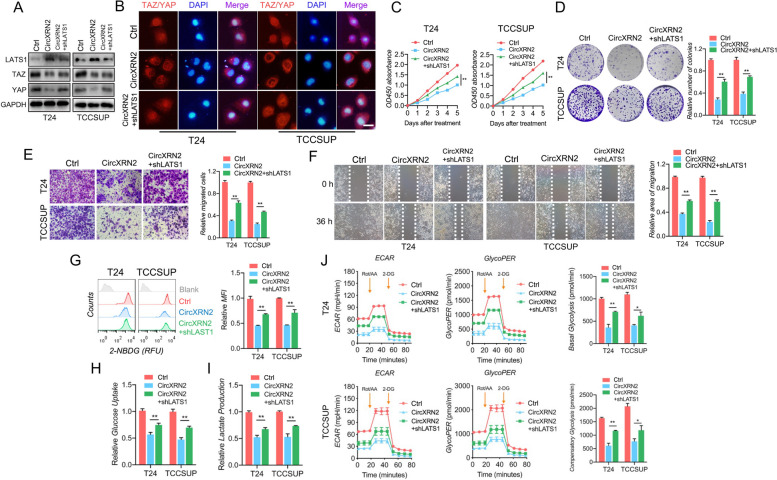
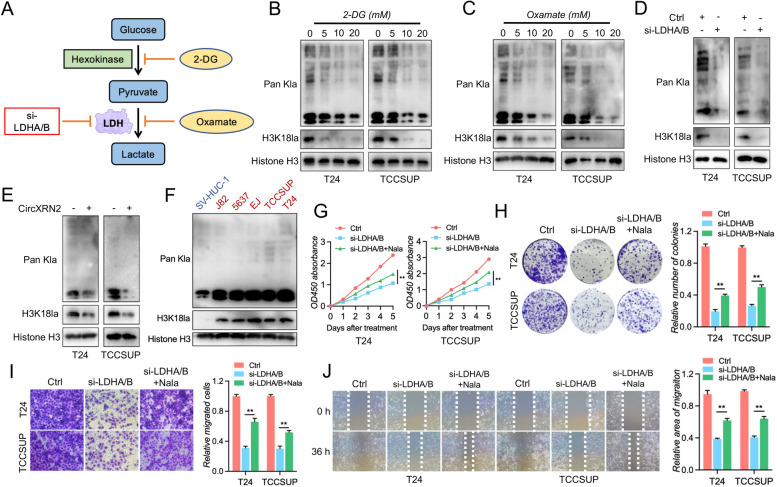
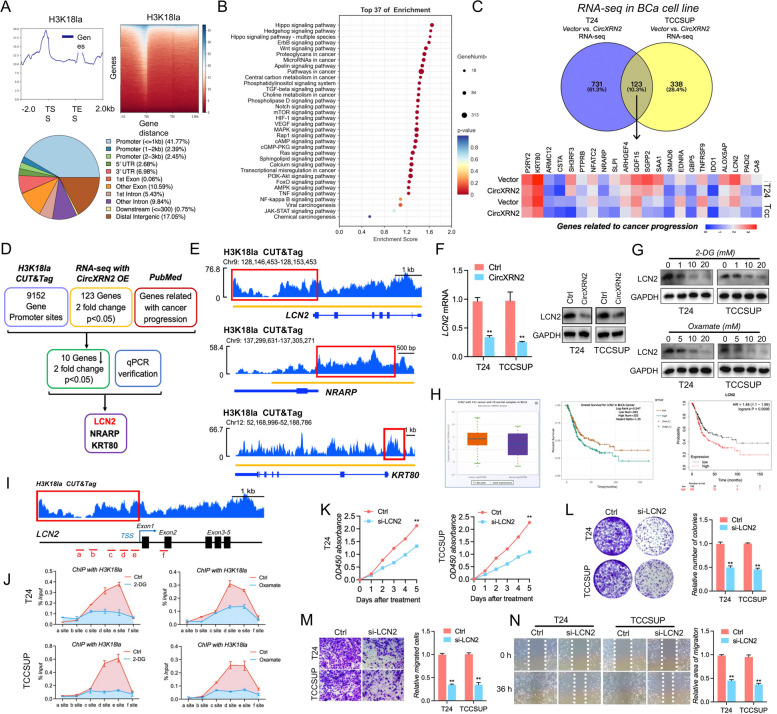
Methods: RNA immunoprecipitation (RIP) followed by circRNA sequencing confirmed circXRN2 as the research object. Overexpression of circXRN2 and knockdown of TAZ/YAP further verified the biological functions in T24 and TCCSUP cells. RIP, immunoprecipitation and coimmunoprecipitation were used to elucidate the interaction between circXRN2 and LATS1. A Seahorse metabolic analyzer was used to determine the glycolytic rate. Cleavage under targets and Tagmentation (CUT&Tag) and chromatin immunoprecipitation (ChIP) were employed to ensure the regulatory roles of H3K18 lactylation in the transcriptional activity of LCN2.
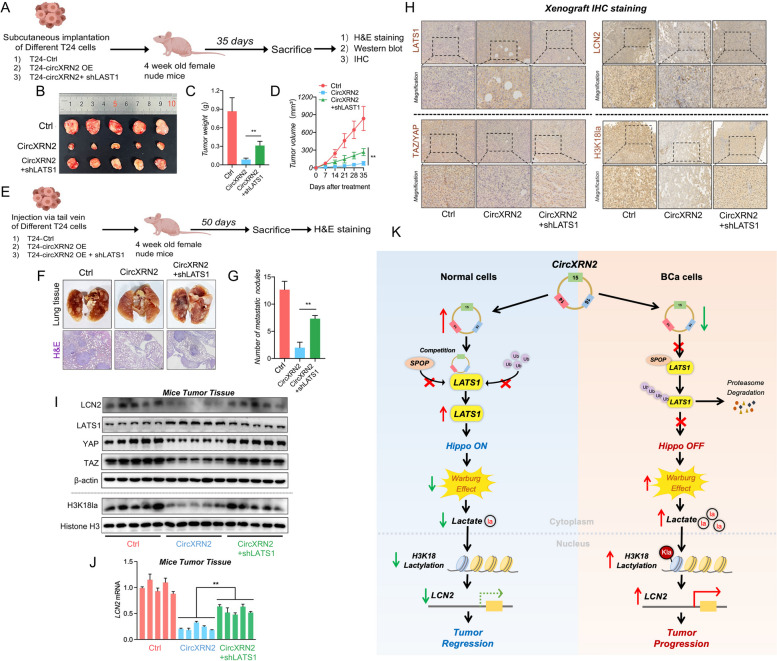
Results: CircXRN2 is aberrantly downregulated in bladder cancer tissues and cell lines. CircXRN2 inhibits the proliferation and migration of tumor cells both in vitro and in vivo. In addition, circXRN2 serves as a negative regulator of glycolysis and lactate production. Mechanistically, circXRN2 prevents LATS1 from SPOP-mediated degradation by binding to the SPOP degron and then activates the Hippo signaling pathway to exert various biological functions. The circXRN2-Hippo pathway regulatory axis further modulates tumor progression by inhibiting H3K18 lactylation and LCN2 expression in human bladder cancer.
Conclusions: CircXRN2 suppresses tumor progression driven by H3K18 lactylation by activating the Hippo signaling pathway in human bladder cancer. Our results indicated novel therapeutic targets and provided promising strategies for clinical intervention in human bladder cancer.
Keywords: Bladder cancer; Hippo pathway; Histone lactylation; Tumor progression; circXRN2.
© 2023. BioMed Central Ltd., part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
