

Lactate-upregulated NADPH-dependent NOX4 expression via HCAR1/PI3K pathway contributes to ROS-induced osteoarthritis chondrocyte damage
- PMID: 37688977
- PMCID: PMC10498433
- DOI: 10.1016/j.redox.2023.102867
Lactate-upregulated NADPH-dependent NOX4 expression via HCAR1/PI3K pathway contributes to ROS-induced osteoarthritis chondrocyte damage
Abstract
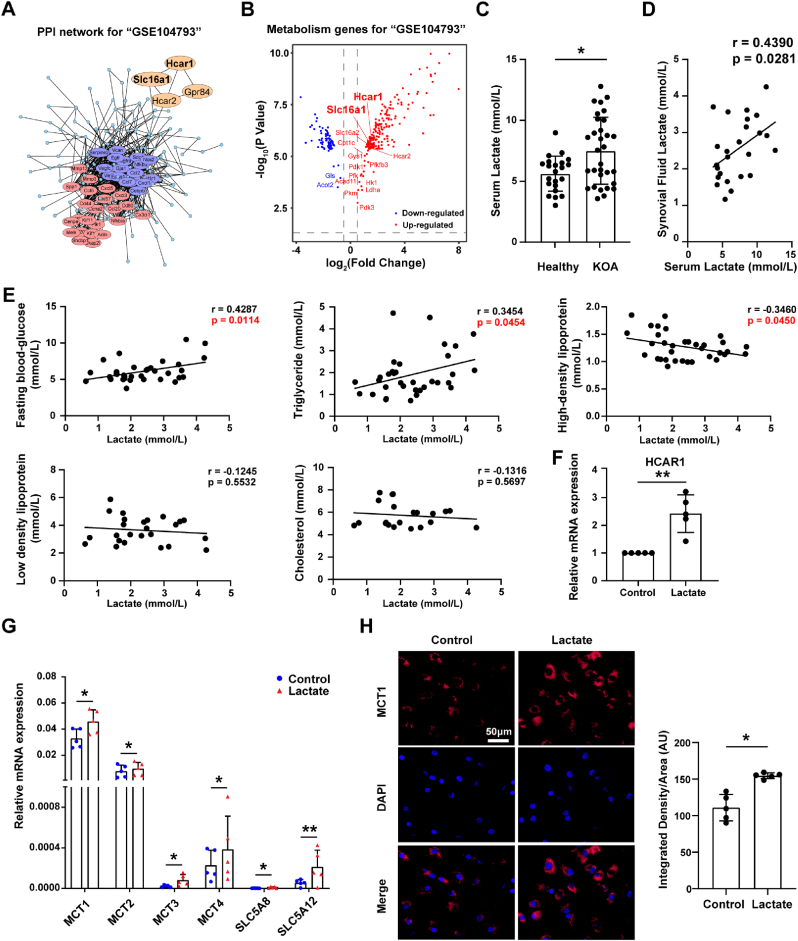
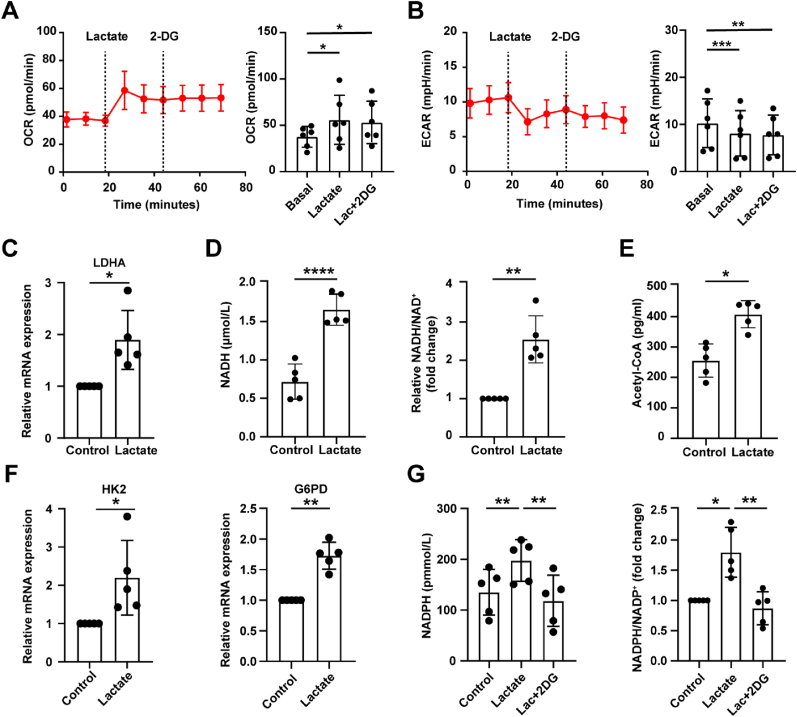
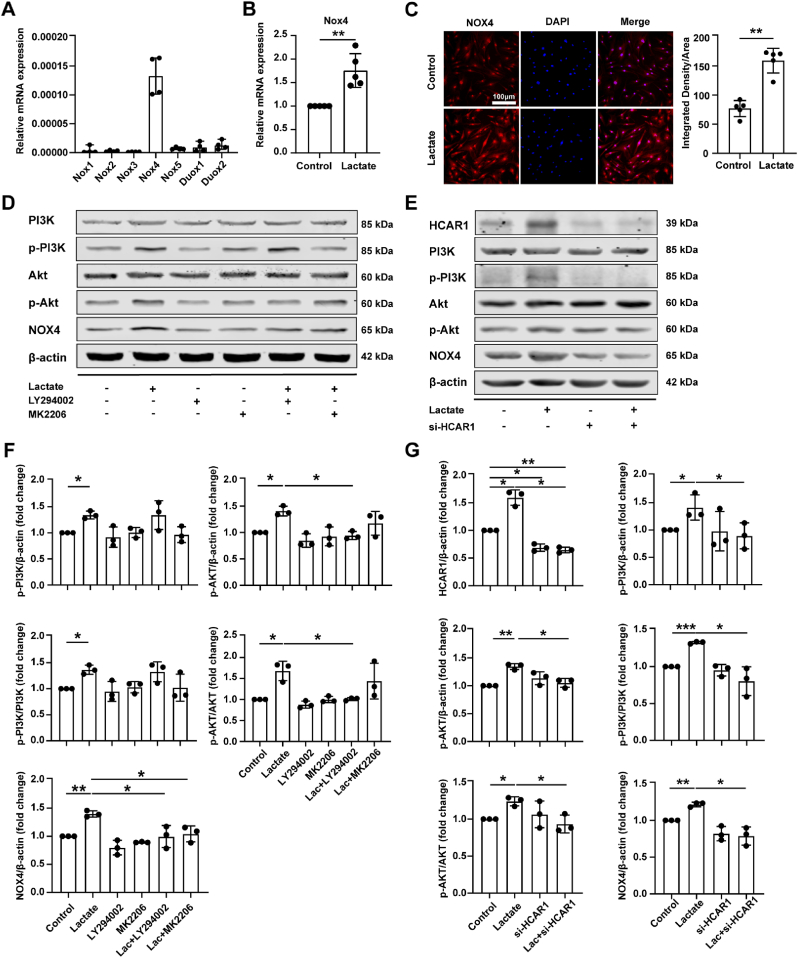
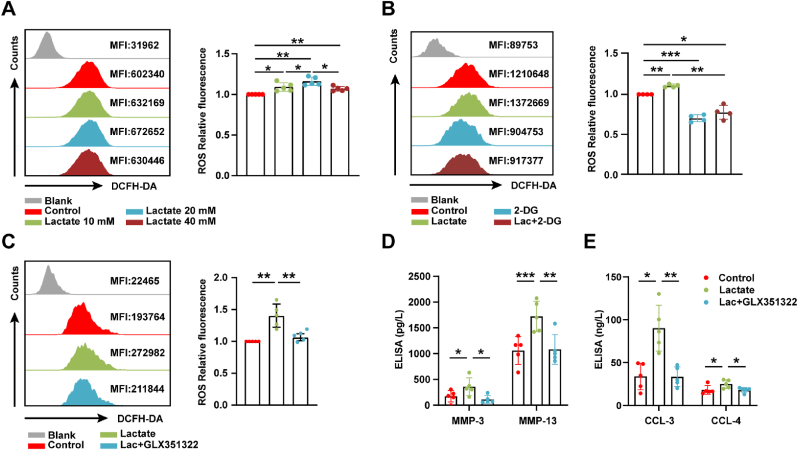
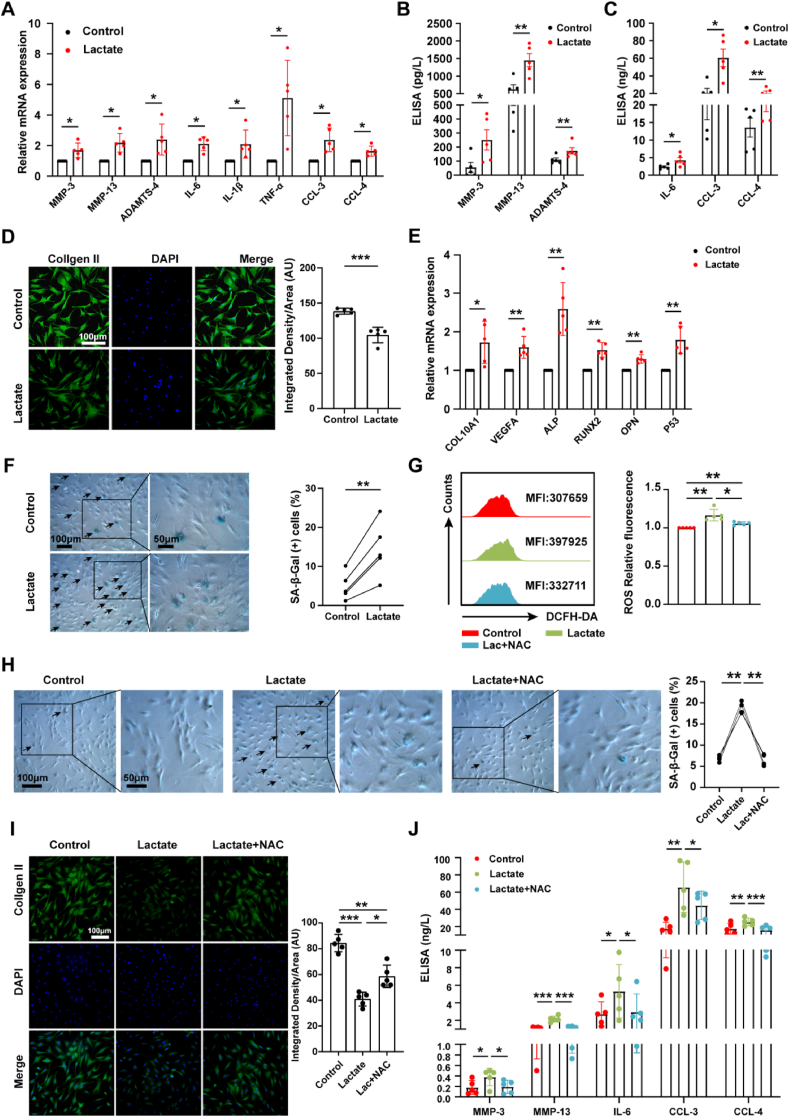
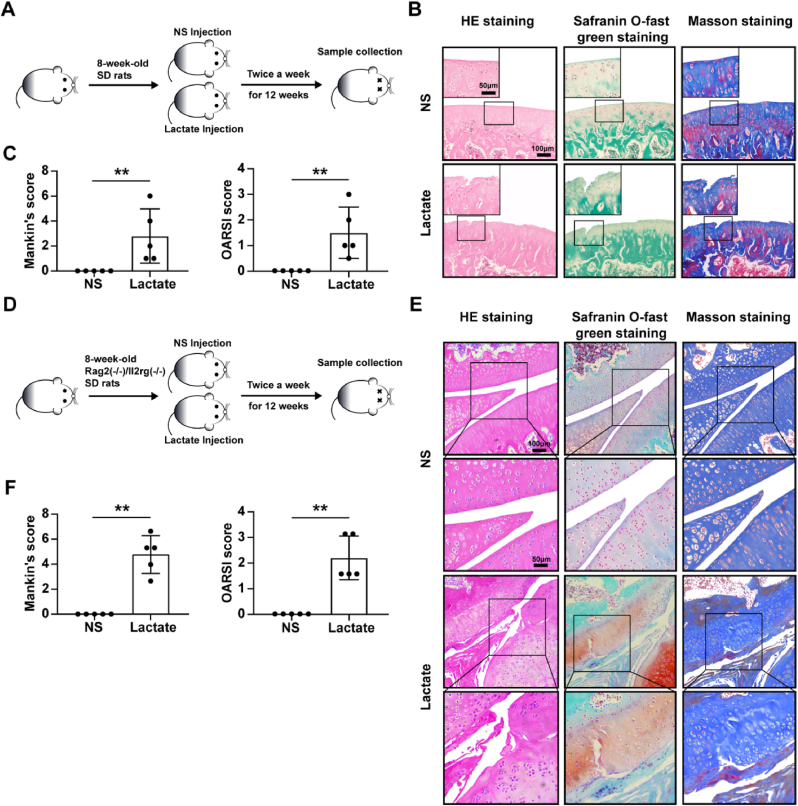
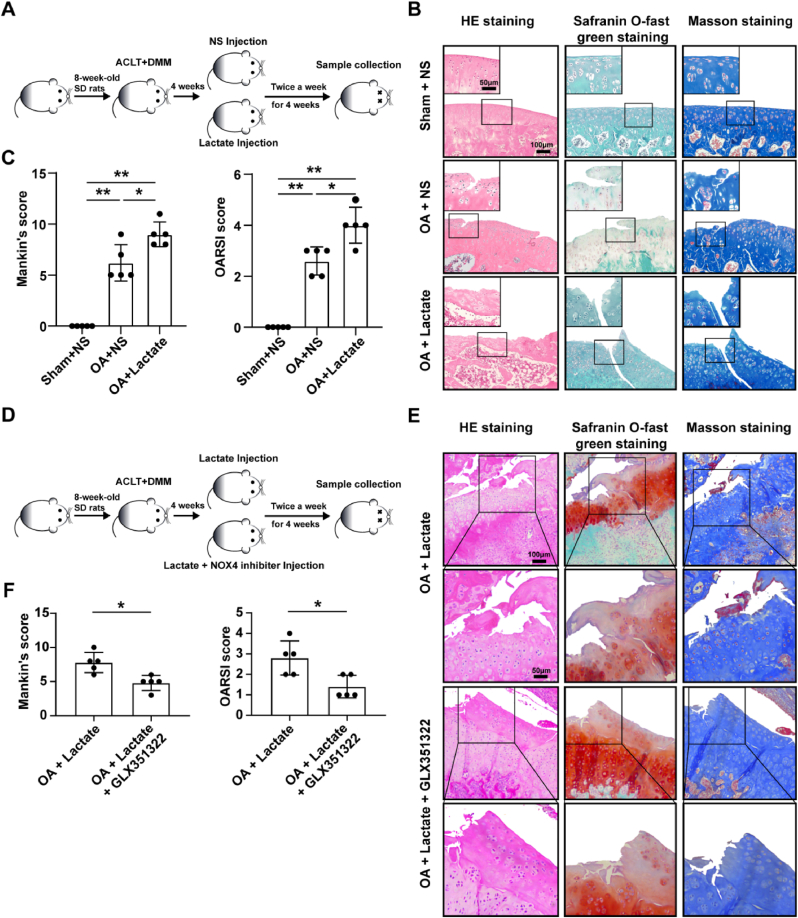
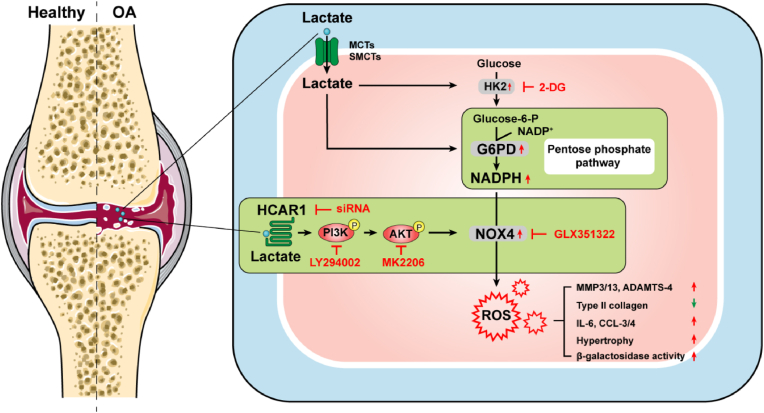
Increasing evidence shows that metabolic factors are involved in the pathological process of osteoarthritis (OA). Lactate has been shown to contribute to the onset and progression of diseases. While whether lactate is involved in the pathogenesis of OA through impaired chondrocyte function and its mechanism remains unclear. This study confirmed that serum lactate levels were elevated in OA patients compared to healthy controls and were positively correlated with synovial fluid lactate levels, which were also correlated with fasting blood glucose, high-density lipoprotein, triglyceride. Lactate treatment could up-regulate expressions of the lactate receptor hydroxy-carboxylic acid receptor 1 (HCAR1) and lactate transporters in human chondrocytes. We demonstrated the dual role of lactate, which as a metabolite increased NADPH levels by shunting glucose metabolism to the pentose phosphate pathway, and as a signaling molecule up-regulated NADPH oxidase 4 (NOX4) via activating PI3K/Akt signaling pathway through receptor HCAR1. Particularly, lactate could promote reactive oxygen species (ROS) generation and chondrocyte damage, which was attenuated by pre-treatment with the NOX4 inhibitor GLX351322. We also confirmed that lactate could increase expression of catabolic enzymes (MMP-3/13, ADAMTS-4), reduce the synthesis of type II collagen, promote expression of inflammatory cytokines (IL-6, CCL-3/4), and induce cellular hypertrophy and aging in chondrocytes. Subsequently, we showed that chondrocyte damage mediated by lactate could be reversed by pre-treatment with N-Acetyl-l-cysteine (NAC, ROS scavenger). Finally, we further verified in vivo that intra-articular injection of lactate in Sprague Dawley (SD) rat models could damage cartilage and exacerbate the progression of OA models that could be countered by the NOX4 inhibitor GLX351322. Our study highlights the involvement of lactate as a metabolic factor in the OA process, providing a theoretical basis for potential metabolic therapies of OA in the future.
Keywords: Chondrocytes; Lactate; NADPH oxidase 4; Osteoarthritis.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper
Figures
References
-
- Cao Y., Tang S., Ding C. Inflammatory phenotype of osteoarthritis and its potential therapies. Rheumatol. Autoimmun. 2021;1:92–100. doi: 10.1002/rai2.12018. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
