

ERG K+ channels mediate a major component of action potential repolarization in lymphatic muscle
- PMID: 37689781
- PMCID: PMC10492848
- DOI: 10.1038/s41598-023-41995-5
ERG K+ channels mediate a major component of action potential repolarization in lymphatic muscle
Abstract
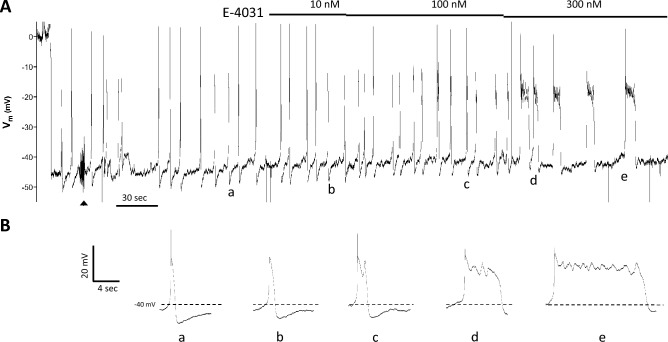
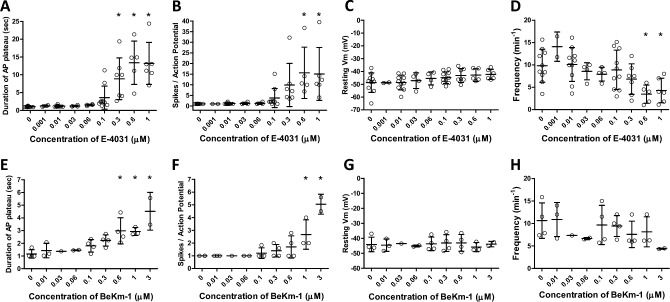
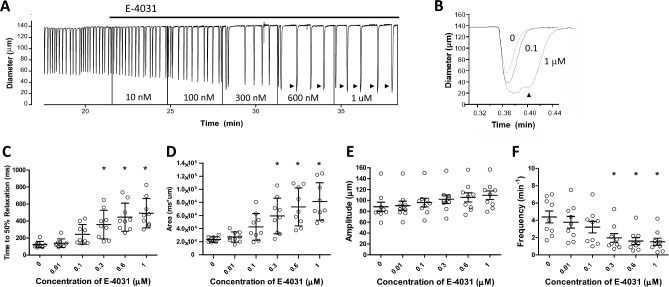
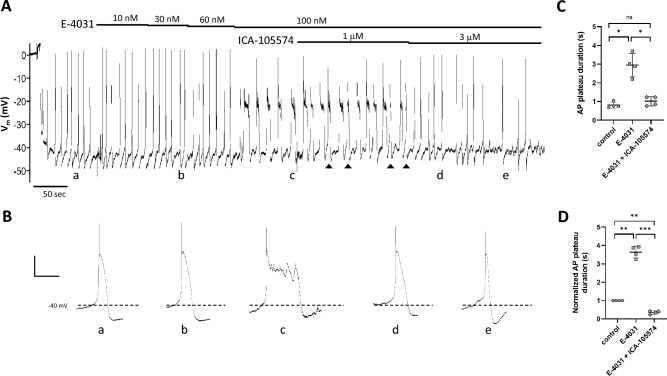
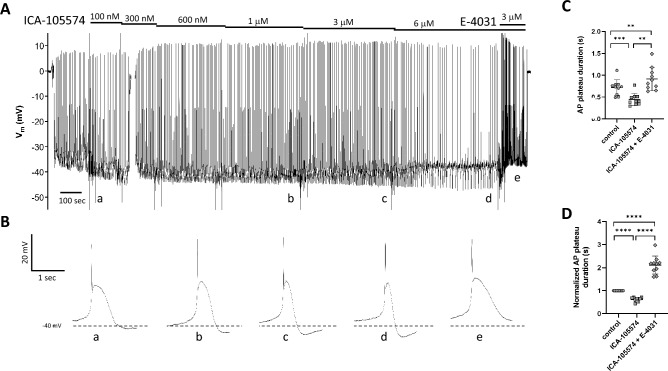
Smooth muscle cells in the walls of collecting lymphatic vessels fire spontaneous action potentials (APs), which conduct rapidly over the muscle layer to initiate contractions that propel lymph. Several ion channels have been implicated in the currents underlying the AP spike and the preceding diastolic depolarization, but the molecular identities of K+ channels involved in AP repolarization are unknown. Based on previous studies of other rhythmically active smooth muscles, we hypothesized that ether-a-go-go related gene (ERG) K+ channels (Kv11) play an important role in repolarization of the AP in lymphatic muscle. Message for one or more ERG channel isoforms was detected by RT-PCR analysis of lymphatic vessels from mice, rats and humans. Membrane potential recordings in smooth muscle cells of rat and human lymphatics revealed that nanomolar concentrations of ERG-1 inhibitors (E-4031 and BeKm-1) prolonged the duration of the AP plateau (normally ~ 1 s in duration) and induced multiple spikes, whereas ERG-1 activators (ICA-105574 and RPR-260243) shortened the plateau and could completely inhibit spontaneous APs. At relatively high inhibitor concentrations, the AP plateau duration lasted as long as 24 s. ERG activators reversed the effects of ERG inhibitors and vice-versa. In pressure myograph studies, ERG channel inhibition prolonged the diastolic repolarization phase of the contraction cycle and reduced the frequency of spontaneous contractions. This is the first evidence for a specific K+ channel contributing to the AP in lymphatic muscle. Our results imply that lymphatic contractile dysfunction may occur in long QT type II patients with mutations that result in ERG channel loss-of-function or impaired trafficking of the channel to the cell membrane.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Olszewski WL, Engeset A. Intrinsic contractility of prenodal lymph vessels and lymph flow in human leg. Am. J. Physiol. 1980;239:H775–783. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
