

Insulin-like growth factor binding protein-3 mediates hyperosmolar stress-induced mitophagy through the mechanistic target of rapamycin
- PMID: 37690686
- PMCID: PMC10637961
- DOI: 10.1016/j.jbc.2023.105239
Insulin-like growth factor binding protein-3 mediates hyperosmolar stress-induced mitophagy through the mechanistic target of rapamycin
Abstract
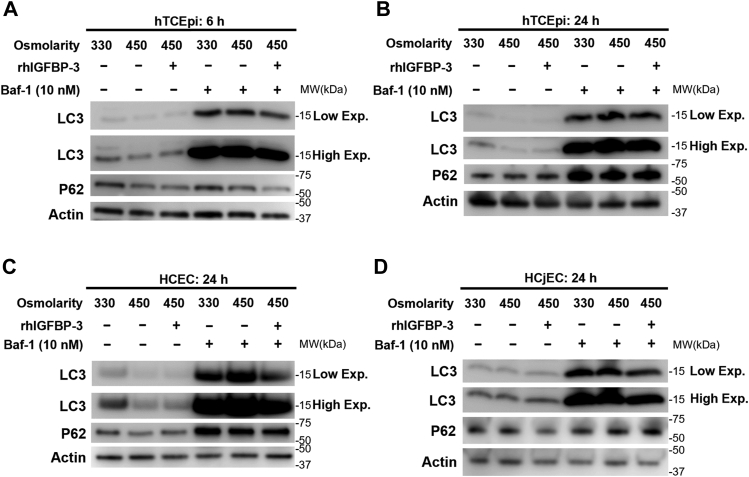
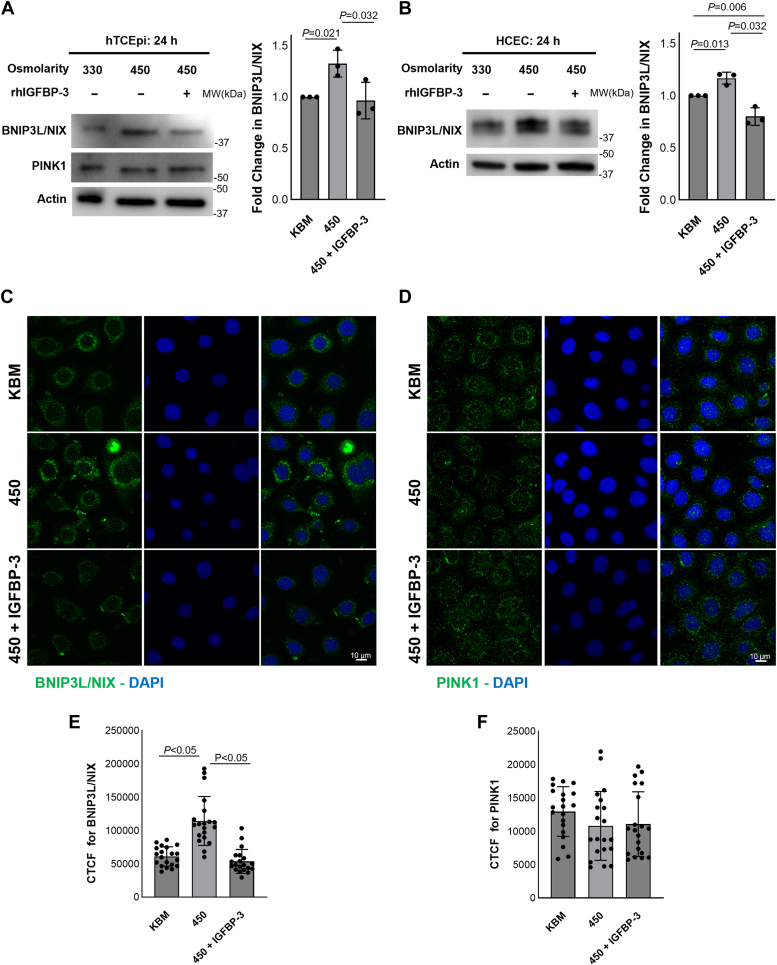
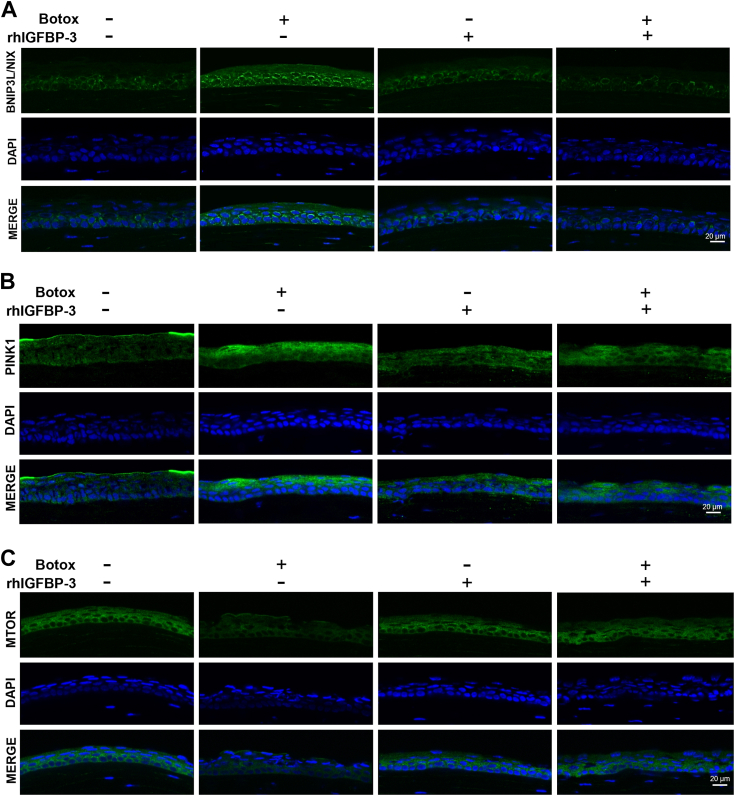
Hyperosmolarity of the ocular surface triggers inflammation and pathological damage in dry eye disease (DED). In addition to a reduction in quality of life, DED causes vision loss and when severe, blindness. Mitochondrial dysfunction occurs as a consequence of hyperosmolar stress. We have previously reported on a role for the insulin-like growth factor binding protein-3 (IGFBP-3) in the regulation of mitochondrial ultrastructure and metabolism in mucosal surface epithelial cells; however, this appears to be context-specific. Due to the finding that IGFBP-3 expression is decreased in response to hyperosmolar stress in vitro and in an animal model of DED, we next sought to determine whether the hyperosmolar stress-mediated decrease in IGFBP-3 alters mitophagy, a key mitochondrial quality control mechanism. Here we show that hyperosmolar stress induces mitophagy through differential regulation of BNIP3L/NIX and PINK1-mediated pathways. In corneal epithelial cells, this was independent of p62. The addition of exogenous IGFBP-3 abrogated the increase in mitophagy. This occurred through regulation of mTOR, highlighting the existence of a new IGFBP-3-mTOR signaling pathway. Together, these findings support a novel role for IGFBP-3 in mediating mitochondrial quality control in DED and have broad implications for epithelial tissues subject to hyperosmolar stress and other mitochondrial diseases.
Keywords: cornea; hyperosmolar stress; mTOR; mitochondria; mitophagy.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures









References
-
- Craig J.P., Nichols K.K., Akpek E.K., Caffery B., Dua H.S., Joo C.K., et al. TFOS DEWS II definition and classification report. Ocul. Surf. 2017;15:276–283. - PubMed
-
- Research in dry eye: report of the research subcommittee of the International dry eye WorkShop (2007) Ocul. Surf. 2007;5:179–193. - PubMed
-
- Png E., Samivelu G.K., Yeo S.H., Chew J., Chaurasia S.S., Tong L. Hyperosmolarity-mediated mitochondrial dysfunction requires transglutaminase-2 in human corneal epithelial cells. J. Cell. Physiol. 2011;226:693–699. - PubMed
-
- Stookey J.D., Barclay D., Arieff A., Popkin B.M. The altered fluid distribution in obesity may reflect plasma hypertonicity. Eur. J. Clin. Nutr. 2007;61:190–199. - PubMed
-
- Vernia P., Gnaedinger A., Hauck W., Breuer R.I. Organic anions and the diarrhea of inflammatory bowel disease. Dig. Dis. Sci. 1988;33:1353–1358. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
