

This is a preprint.
Repair of CRISPR-guided RNA breaks enables site-specific RNA editing in human cells
- PMID: 37693568
- PMCID: PMC10491232
- DOI: 10.1101/2023.08.29.555404
Repair of CRISPR-guided RNA breaks enables site-specific RNA editing in human cells
Update in
-
Repair of CRISPR-guided RNA breaks enables site-specific RNA excision in human cells.Science. 2024 May 17;384(6697):808-814. doi: 10.1126/science.adk5518. Epub 2024 Apr 25. Science. 2024. PMID: 38662916 Free PMC article.
Abstract
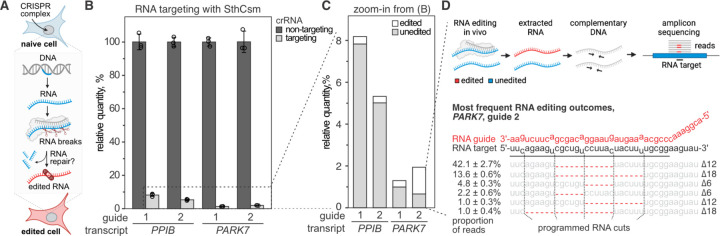
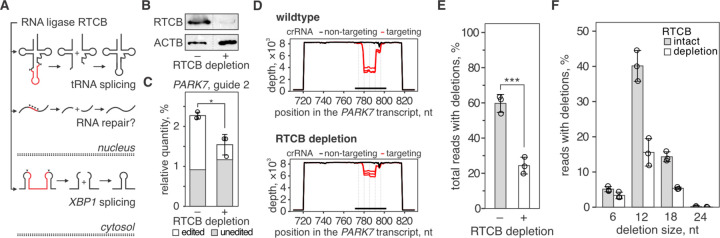
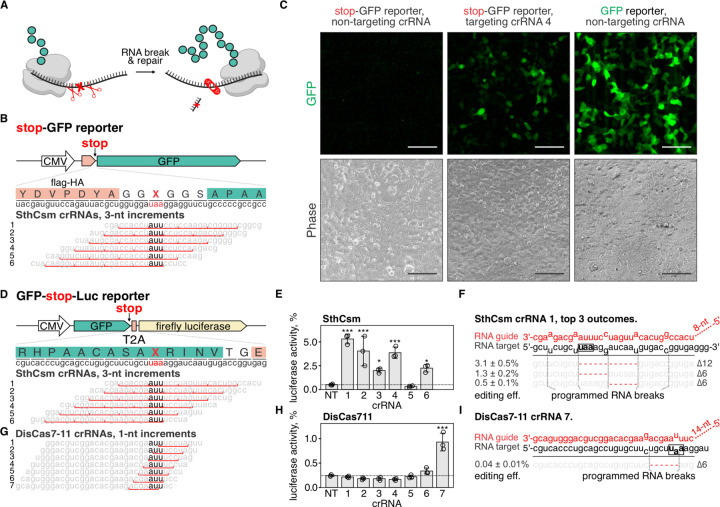
Genome editing with CRISPR RNA-guided endonucleases generates DNA breaks that are resolved by cellular DNA repair machinery. However, analogous methods to manipulate RNA remain unavailable. Here, we show that site-specific RNA breaks generated with RNA-targeting CRISPR complexes are repaired in human cells, and this repair can be used for programmable deletions in human transcripts that restore gene function. Collectively, this work establishes a technology for precise RNA manipulation with potential therapeutic applications.
Conflict of interest statement
Competing interests: B.W. is the founder of SurGene LLC and VIRIS Detection Systems Inc. B.W., A. Nemudryi, and A. Nemudraia are inventors of the patent applications US 63/523,592 and US 63/534,305 pertaining to use type III CRISPR-Cas system for sequence-specific editing of RNA filed by Montana State University.
Figures
References
-
- van Overbeek M. et al. , DNA repair profiling reveals nonrandom outcomes at Cas9-mediated Breaks. Mol Cell 63, 633–646 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials