

Two Notorious Nodes: A Critical Examination of Relaxed Molecular Clock Age Estimates of the Bilaterian Animals and Placental Mammals
- PMID: 37695319
- PMCID: PMC11129587
- DOI: 10.1093/sysbio/syad057
Two Notorious Nodes: A Critical Examination of Relaxed Molecular Clock Age Estimates of the Bilaterian Animals and Placental Mammals
Abstract
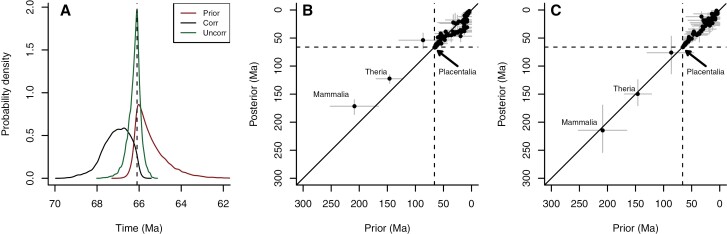
The popularity of relaxed clock Bayesian inference of clade origin timings has generated several recent publications with focal results considerably older than the fossils of the clades in question. Here, we critically examine two such clades: the animals (with a focus on the bilaterians) and the mammals (with a focus on the placentals). Each example displays a set of characteristic pathologies which, although much commented on, are rarely corrected for. We conclude that in neither case does the molecular clock analysis provide any evidence for an origin of the clade deeper than what is suggested by the fossil record. In addition, both these clades have other features (including, in the case of the placental mammals, proximity to a large mass extinction) that allow us to generate precise expectations of the timings of their origins. Thus, in these instances, the fossil record can provide a powerful test of molecular clock methodology, and why it goes astray, and we have every reason to think these problems are general. [Cambrian explosion; mammalian evolution; molecular clocks.].
© The Author(s) 2023. Published by Oxford University Press on behalf of the Society of Systematic Biologists.
Conflict of interest statement
The authors have no competing interests to declare.
Figures





References
-
- Alroy J. 1999. The fossil record of North American mammals: evidence for a paleocene evolutionary radiation. Syst. Biol. 48:107–118. - PubMed
-
- Álvarez-Carretero S., Tamuri A.U., Battini M., Nascimento F.F., Carlisle E., Asher R.J., Yang Z., Donoghue P.C.J., dos Reis M.. 2022. A species-level timeline of mammal evolution integrating phylogenomic data. Nature 602:263–267. - PubMed
