

CD8+ tissue-resident memory T-cell development depends on infection-matching regulatory T-cell types
- PMID: 37696824
- PMCID: PMC10495327
- DOI: 10.1038/s41467-023-41364-w
CD8+ tissue-resident memory T-cell development depends on infection-matching regulatory T-cell types
Abstract
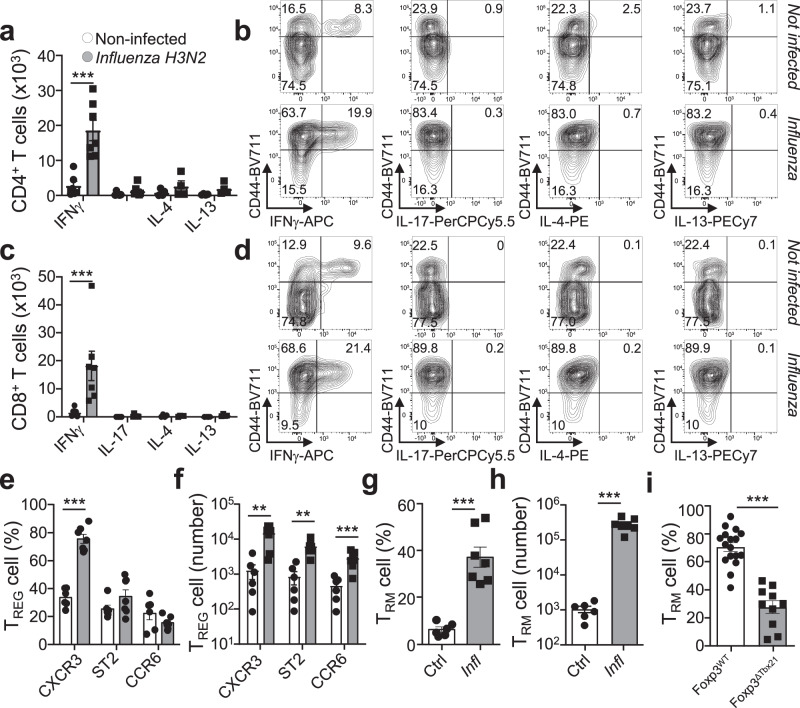
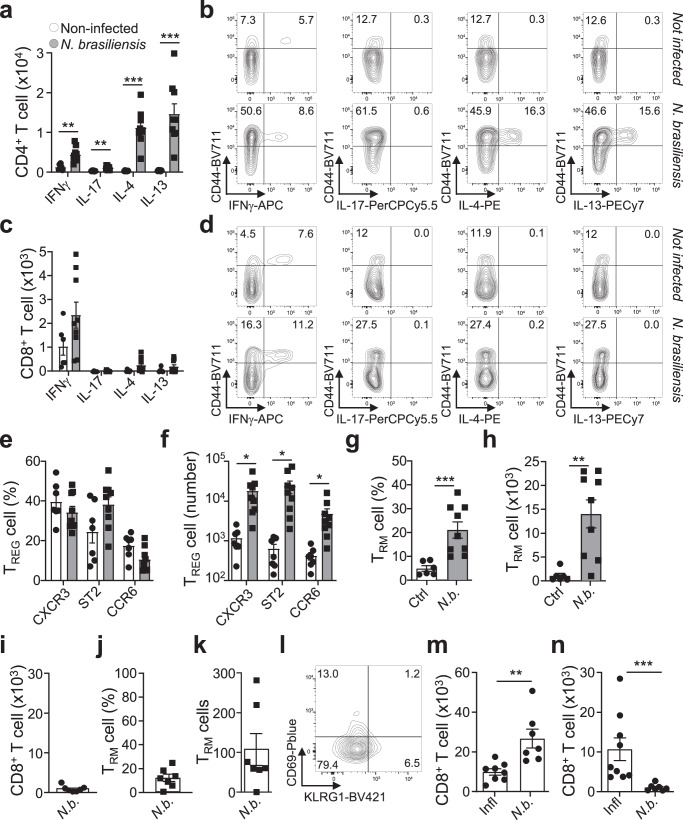
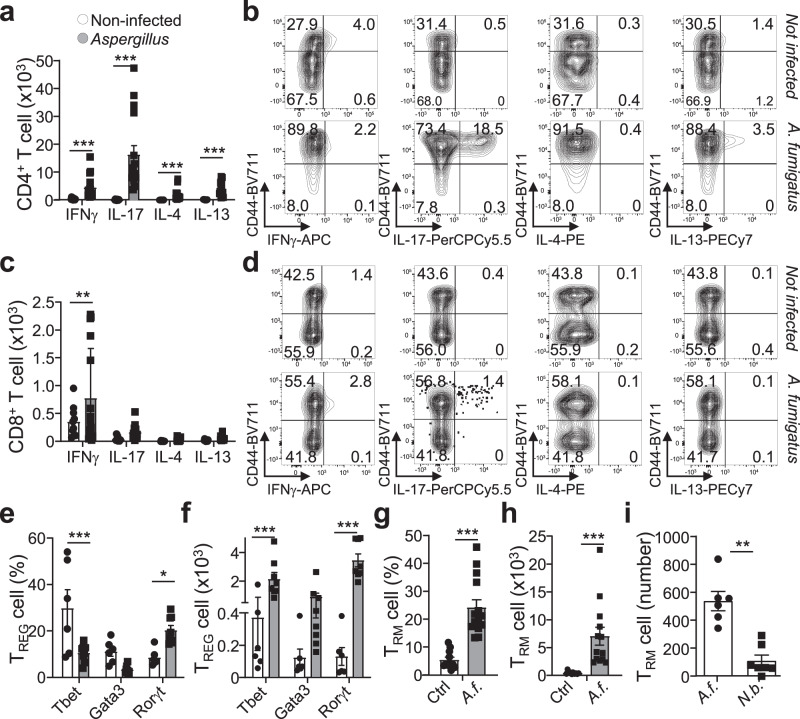
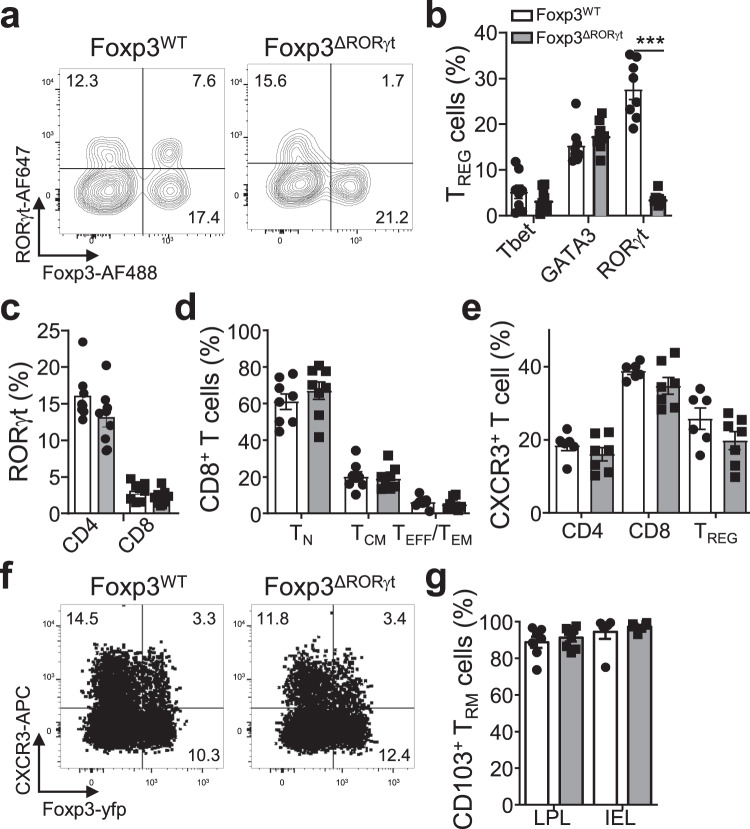
Immunological memory is critical for immune protection, particularly at epithelial sites, which are under constant risk of pathogen invasions. To counter invading pathogens, CD8+ memory T cells develop at the location of infection: tissue-resident memory T cells (TRM). CD8+ T-cell responses are associated with type-1 infections and type-1 regulatory T cells (TREG) are important for CD8+ T-cell development, however, if CD8+ TRM cells develop under other infection types and require immune type-specific TREG cells is unknown. We used three distinct lung infection models, to show that type-2 helminth infection does not establish CD8+ TRM cells. Intracellular (type-1) and extracellular (type-3) infections do and rely on the recruitment of response type-matching TREG population contributing transforming growth factor-β. Nevertheless, type-1 TREG cells remain the most important population for TRM cell development. Once established, TRM cells maintain their immune type profile. These results may have implications in the development of vaccines inducing CD8+ TRM cells.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Masopust D, Vezys V, Marzo AL, Lefrancois L. Preferential localization of effector memory cells in nonlymphoid tissue. Science. 2001;291:2413–2417. - PubMed
-
- Reinhardt RL, Khoruts A, Merica R, Zell T, Jenkins MK. Visualizing the generation of memory CD4 T cells in the whole body. Nature. 2001;410:101–105. - PubMed
-
- Konjar S, Ficht X, Iannacone M, Veldhoen M. Heterogeneity of tissue resident memory T cells. Immunol. Lett. 2022;245:1–7. - PubMed
-
- Shiow LR, et al. CD69 acts downstream of interferon-alpha/beta to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature. 2006;440:540–544. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
