

cloneRate: fast estimation of single-cell clonal dynamics using coalescent theory
- PMID: 37699006
- PMCID: PMC10534056
- DOI: 10.1093/bioinformatics/btad561
cloneRate: fast estimation of single-cell clonal dynamics using coalescent theory
Abstract
Motivation: While evolutionary approaches to medicine show promise, measuring evolution itself is difficult due to experimental constraints and the dynamic nature of body systems. In cancer evolution, continuous observation of clonal architecture is impossible, and longitudinal samples from multiple timepoints are rare. Increasingly available DNA sequencing datasets at single-cell resolution enable the reconstruction of past evolution using mutational history, allowing for a better understanding of dynamics prior to detectable disease. There is an unmet need for an accurate, fast, and easy-to-use method to quantify clone growth dynamics from these datasets.
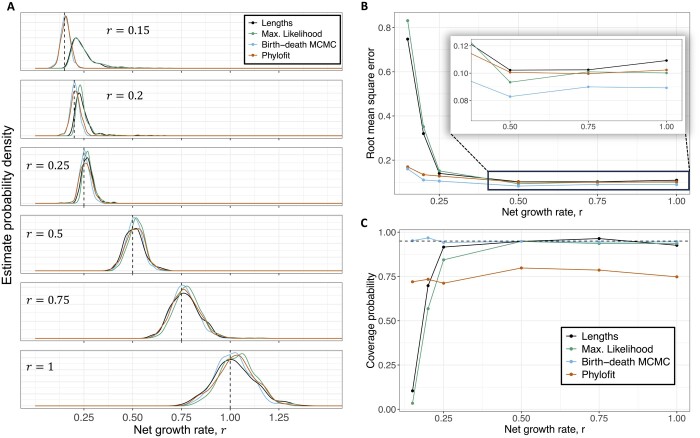
Results: We derived methods based on coalescent theory for estimating the net growth rate of clones using either reconstructed phylogenies or the number of shared mutations. We applied and validated our analytical methods for estimating the net growth rate of clones, eliminating the need for complex simulations used in previous methods. When applied to hematopoietic data, we show that our estimates may have broad applications to improve mechanistic understanding and prognostic ability. Compared to clones with a single or unknown driver mutation, clones with multiple drivers have significantly increased growth rates (median 0.94 versus 0.25 per year; P = 1.6×10-6). Further, stratifying patients with a myeloproliferative neoplasm (MPN) by the growth rate of their fittest clone shows that higher growth rates are associated with shorter time to MPN diagnosis (median 13.9 versus 26.4 months; P = 0.0026).
Availability and implementation: We developed a publicly available R package, cloneRate, to implement our methods (Package website: https://bdj34.github.io/cloneRate/). Source code: https://github.com/bdj34/cloneRate/.
© The Author(s) 2023. Published by Oxford University Press.
Conflict of interest statement
None declared.
Figures





References
-
- Antle C, Klimko L, Harkness W. Confidence intervals for the parameters of the logistic distribution. Biometrika 1970;57:397–402.
