

Death-seq identifies regulators of cell death and senolytic therapies
- PMID: 37699398
- PMCID: PMC10597643
- DOI: 10.1016/j.cmet.2023.08.008
Death-seq identifies regulators of cell death and senolytic therapies
Abstract
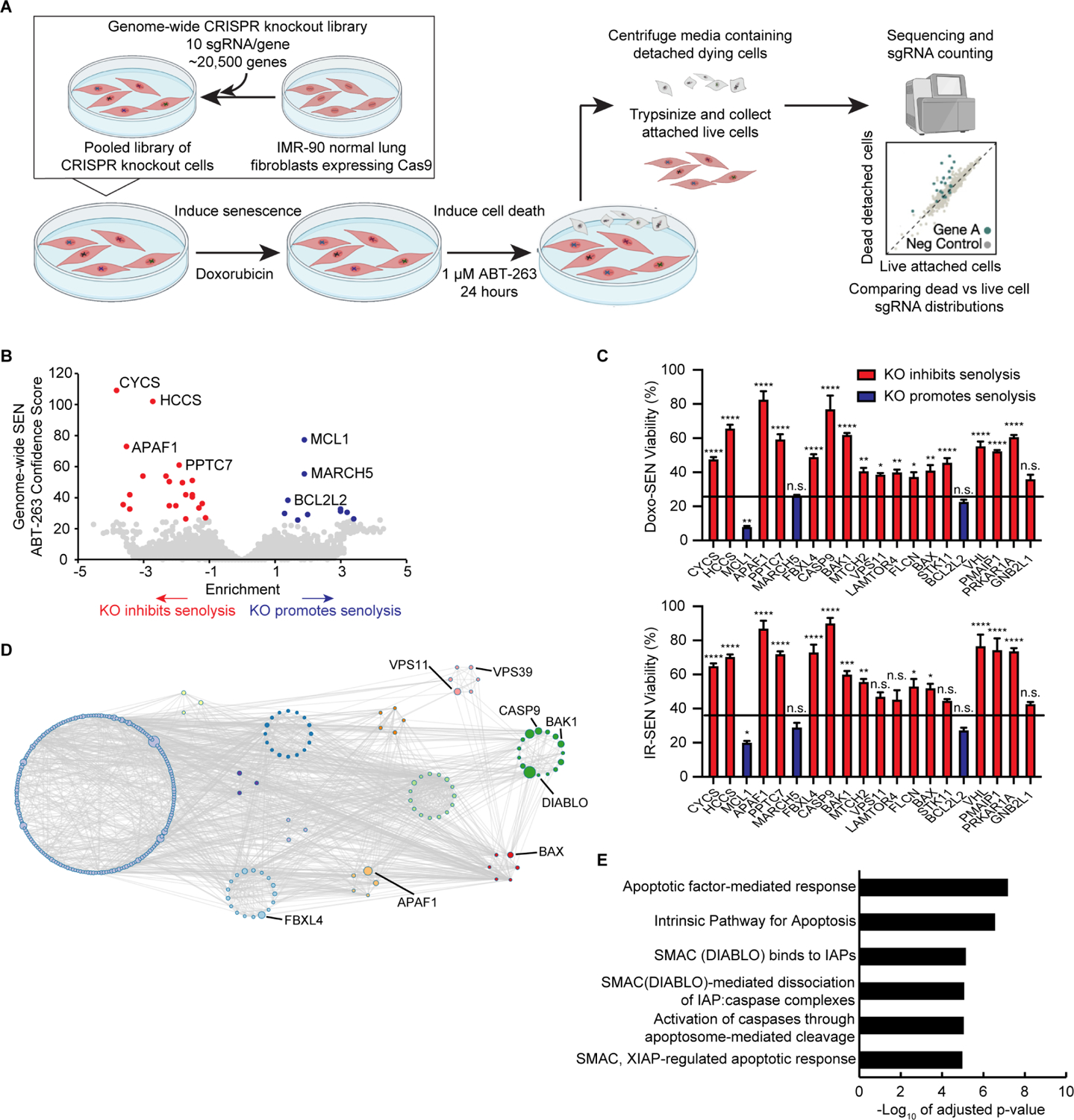
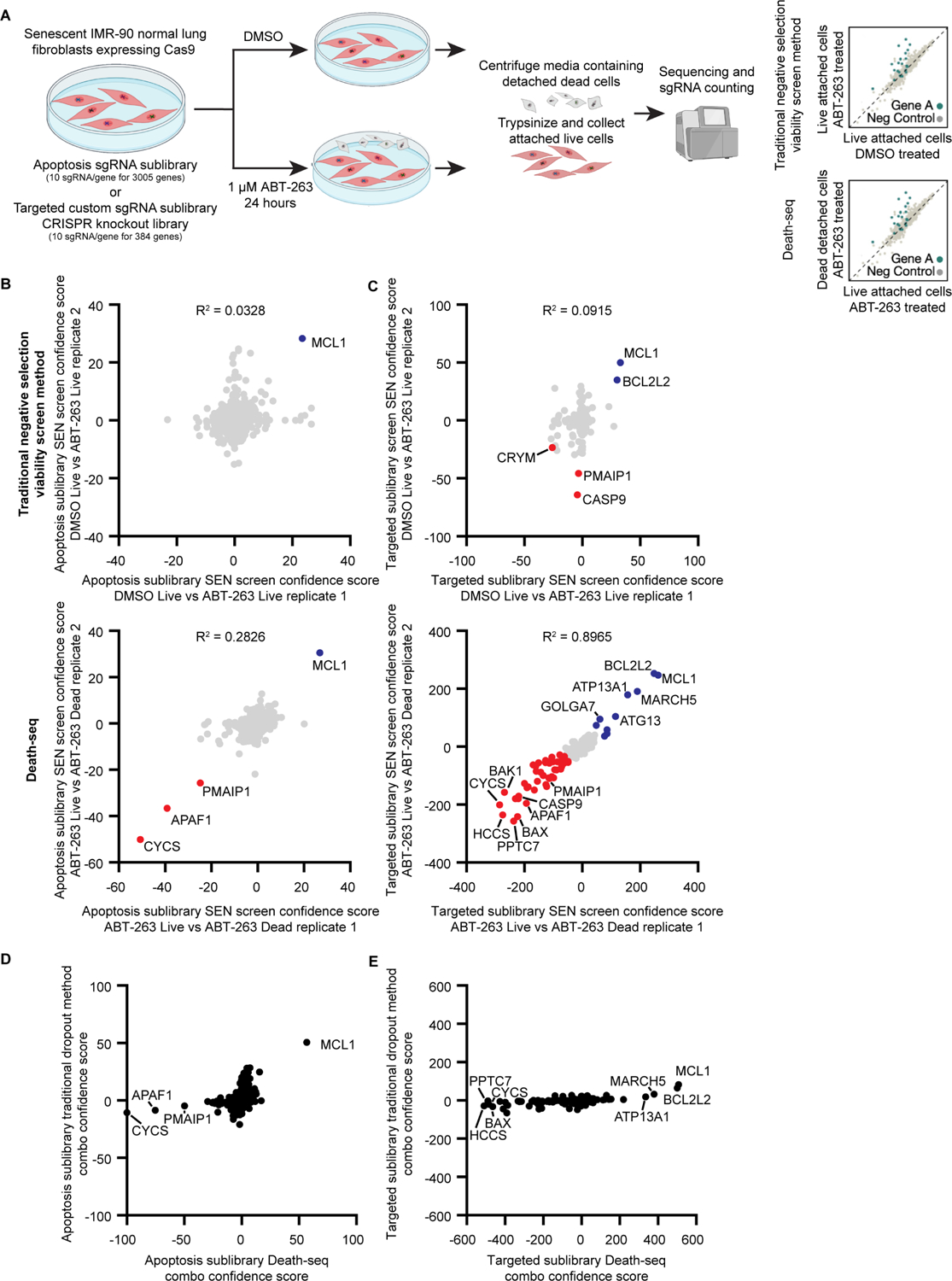
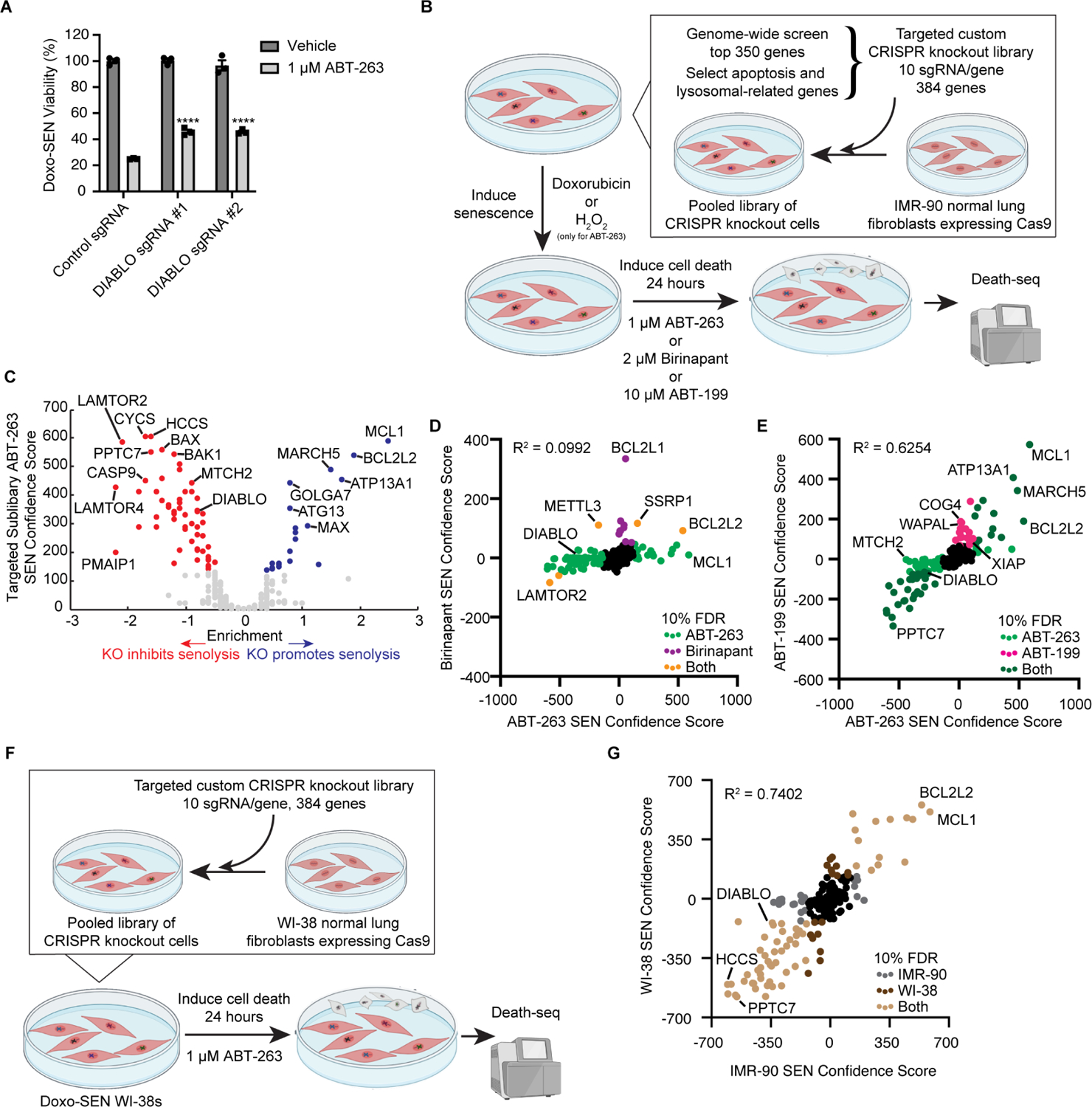
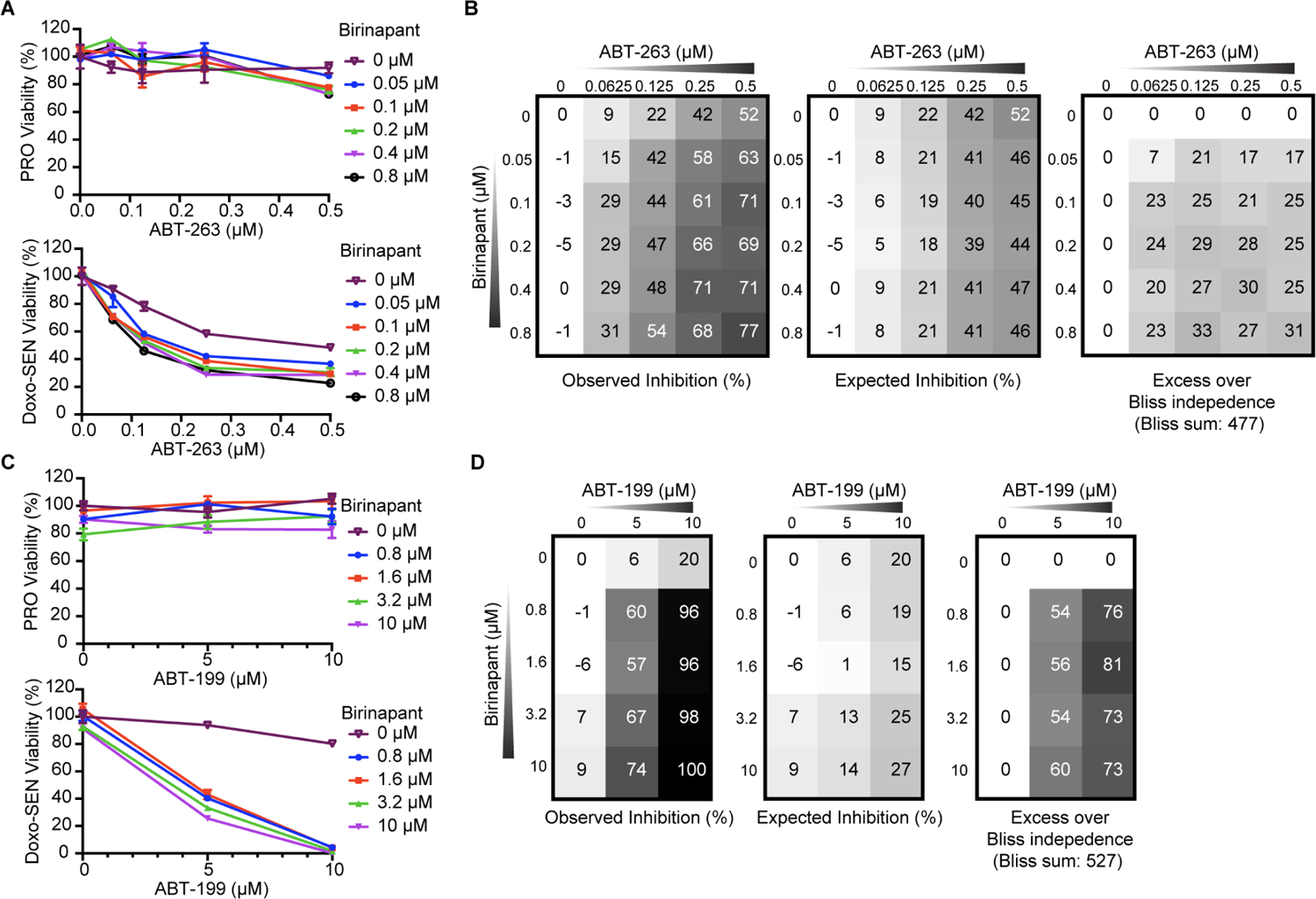
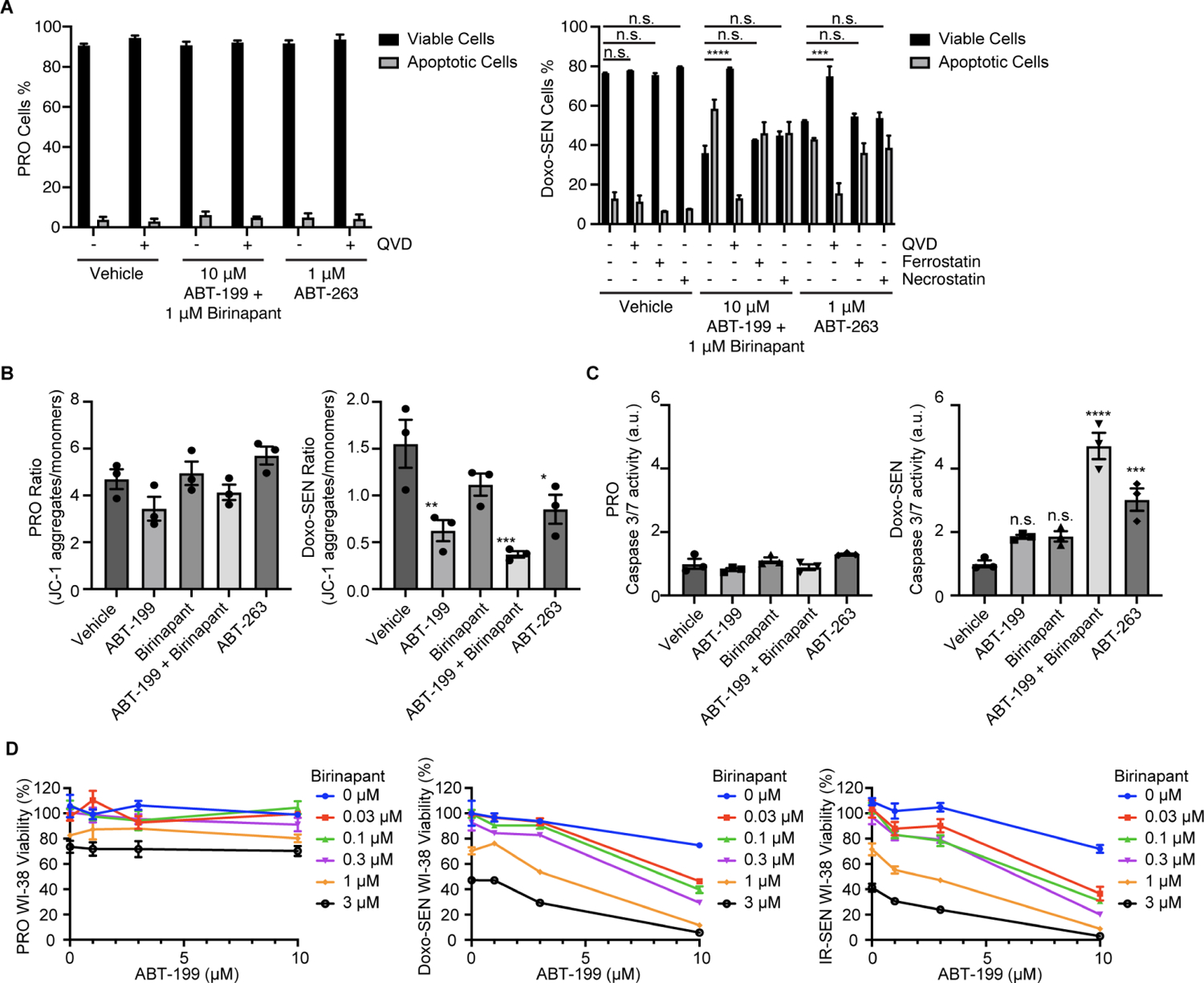
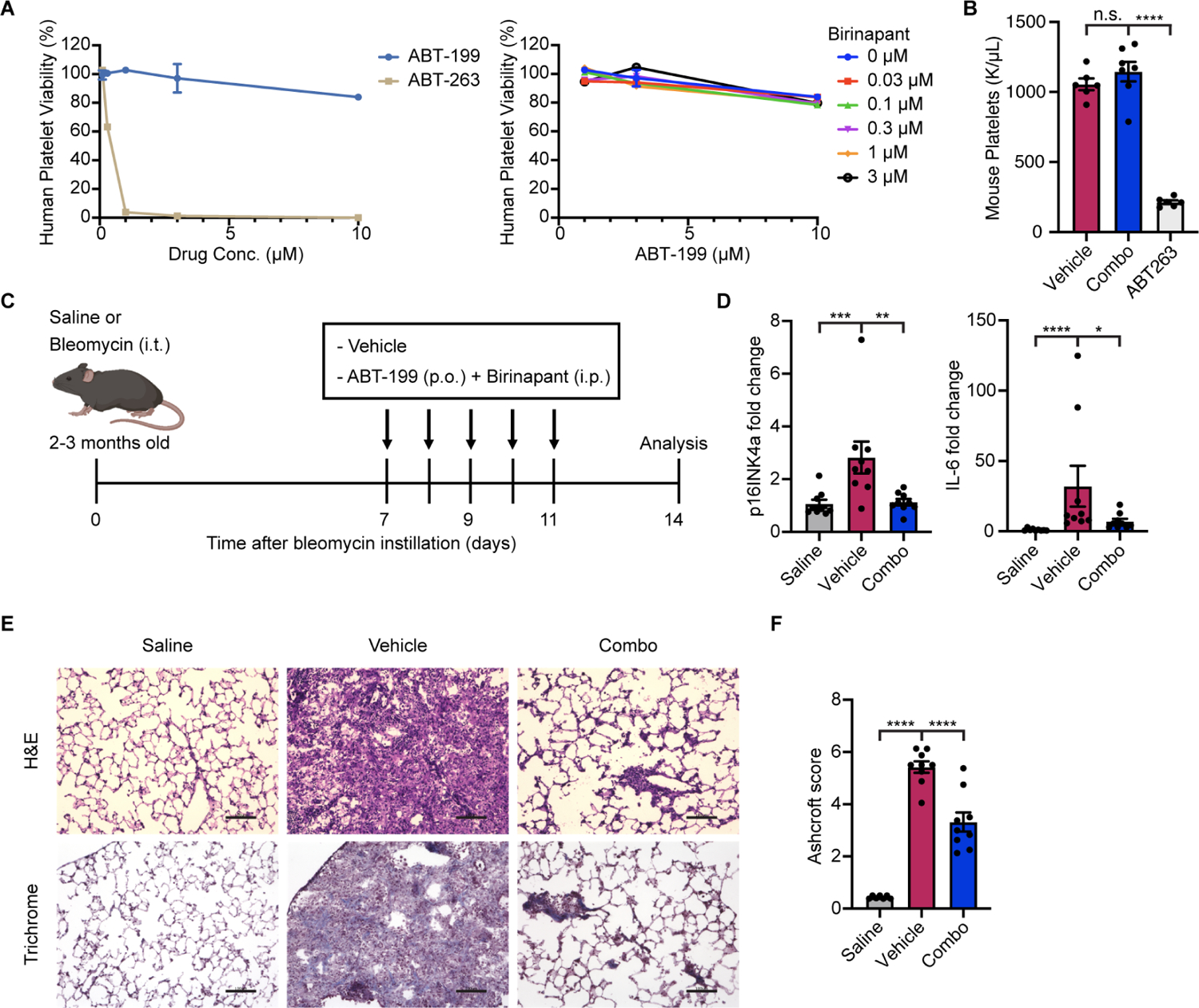
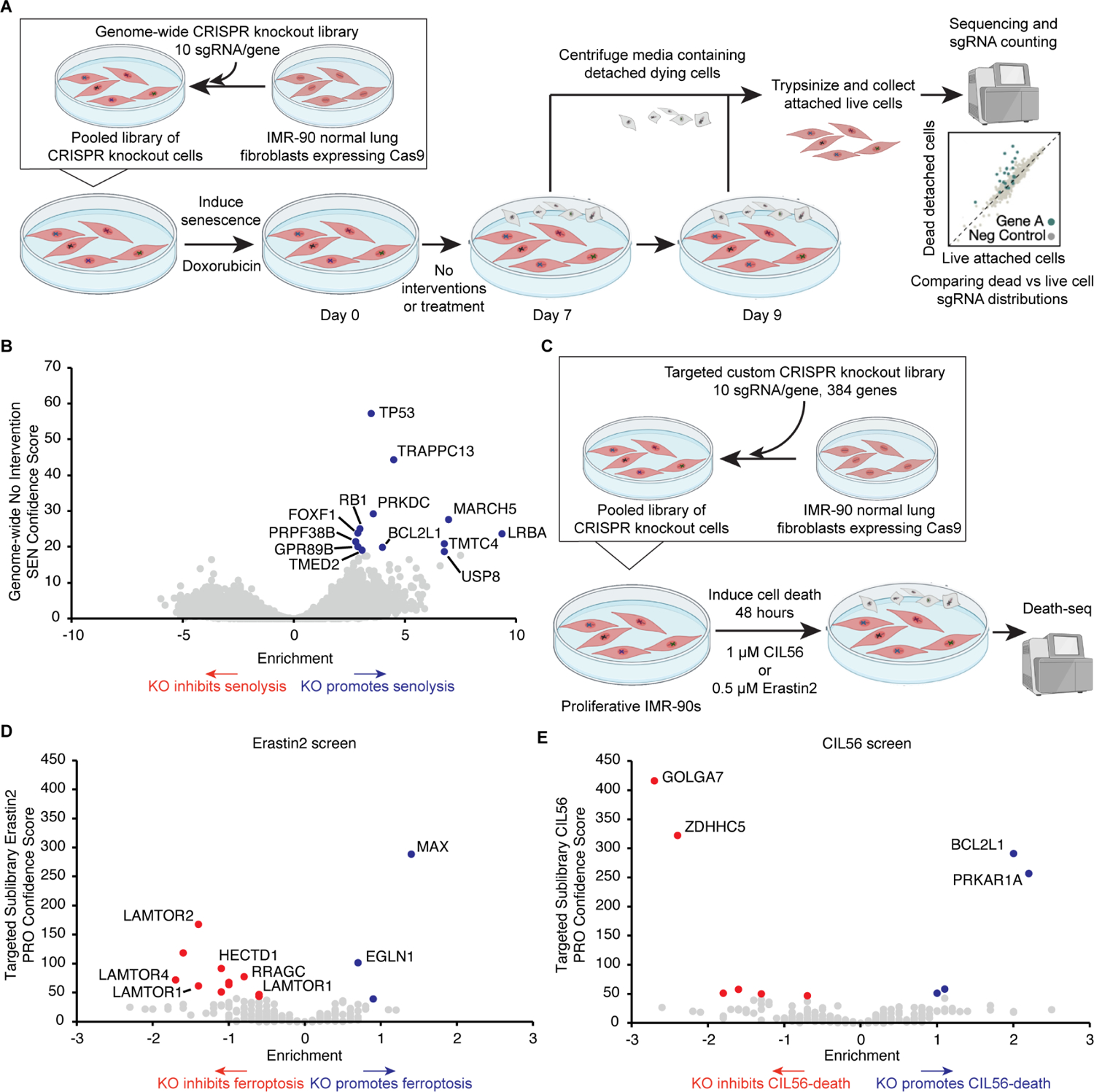
Selectively ablating damaged cells is an evolving therapeutic approach for age-related disease. Current methods for genome-wide screens to identify genes whose deletion might promote the death of damaged or senescent cells are generally underpowered because of the short timescales of cell death as well as the difficulty of scaling non-dividing cells. Here, we establish "Death-seq," a positive-selection CRISPR screen optimized to identify enhancers and mechanisms of cell death. Our screens identified synergistic enhancers of cell death induced by the known senolytic ABT-263. The screen also identified inducers of cell death and senescent cell clearance in models of age-related diseases by a related compound, ABT-199, which alone is not senolytic but exhibits less toxicity than ABT-263. Death-seq enables the systematic screening of cell death pathways to uncover molecular mechanisms of regulated cell death subroutines and identifies drug targets for the treatment of diverse pathological states such as senescence, cancer, and fibrosis.
Keywords: CRISPR; Death-seq; cell death; death screen; genome-wide; positive selection; pulmonary fibrosis; senescence; senolytics; synthetic lethality.
Copyright © 2023 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.C. and T.A.R. have filed a patent application related to the subject matter of this paper. A.C. was formerly a paid consultant during this work for Maze Therapeutics and Rubedo Life Sciences.
Figures
Comment in
-
Death-seq and ye shall find: A novel screening strategy for dying cells.Cell Metab. 2023 Oct 3;35(10):1675-1676. doi: 10.1016/j.cmet.2023.09.003. Cell Metab. 2023. PMID: 37793342
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
